Выбор читателей




Популярные статьи
Функции поджелудочной железы .
I. Экзокринная. Она заключается в секреции панкреатического сока – смеси пищеварительных ферментов, поступающих в двенадцатиперстную кишку и расщепляющих все компоненты химуса;
II. Эндокринная. Она заключается в продукции гормонов.
Поджелудочная железа – паренхиматозный дольчатый орган.
Строма железы представлена: капсулой, которая сливается с висцеральной брюшиной и отходя-щими от неё трабекулами. Строма тонкая, образована рыхлой волокнистой - тканью. Трабекулы делят железу на дольки. В прослойках рыхлой волокнистой ткани находятся выводные протоки экзокринной части железы, сосуды, нервы, интраму-ральные ганглии, пластинчатые тельца Фатер-Пачини.
Паренхима образо-вана совокупностью секреторных отделов (ацинусов ), выводных протоков и остров-ков Лангерганса. Каждая долька состоит из экзокринной и эндокринной частей. Их соотношение ≈ 97: 3.
Экзокринная часть поджелудочной железы представляет собой сложную алъвеолярно-трубчатую белковую железу. Структурно-функциональной единицей экзокринной части является панкреатический ацинус. Он образован 8 – 14 ацинозными клетками (ациноцитами ) и центроацинозными клетками (центроациноцитами ). Ацинозные клетки лежат на базальной мембране, имеют коническую форму и выраженную полярность: различающиеся по строению базальный и апикальный полюсы. Расширенный базальный полюс равномерно окрашивается основными красителями и называется гомогенным. Суженный апикальный полюс окрашивается кислыми красителями и называется зимогенным , потому что содержит гранулы зимогена – проферментов. На апикальном полюсе ациноцитов имеются микроворсинки. Функция ациноцитов – выработка пищеварительных ферментов. Активация ферментов, секретируемых ациноцитами, в норме происходит только в двенадцатиперстной кишке под влиянием активаторов. Это обстоятельство, а также вырабатываемые клетками эпителия протоков ингибиторы ферментов и слизь защищают паренхиму поджелудочной железы от аутолиза (самопереваривания).

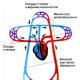
Поджелудочная железа, долька , рисунок, большое увеличение:
1 – концевой отдел (ацинус):
а – апикальная (оксифильная) часть клетки, содержит зимоген,
б – базальная (базофильная) – гомогенная часть клетки;
2 – гемокапилляр;
3 – островок Лангерганса (инсула).
Эндокринная часть железы. Структурно-функциональной единицей эндокринной части поджелудочной железы является островок Лангер ганса (инсула). Он отделён от ацинусов рыхлой волокнистой неоформленной тканью. Островок состоит из клеток инсулоцитов , между которыми лежит рыхлая волокнистая соединительная ткань с гемокапиллярами фенестрированного типа. Инсулоциты различаются по способности окрашиваться красителями. В соответствии с этим различают инсулоциты типа А, В, D, D1, PP.
В-клетки (базофильные инсулоциты) окрашиваются в синий цвет основными красителями. Их количество составляет около 75% всех клеток островка. Они располагаются в центре инсулы. Клетки имеют развитый белоксинтезирующий аппарат и секреторные гранулы с широким светлым ободком. Секреторные гранулы содержат гормон инсулин в комплексе с цинком. Функцией В-инсулоцитов является выработка инсулина, снижающего в крови уровень глюкозы и стимулирующего ее поглощение клетками организма. В печени инсулин стимулирует образование из глюкозы гликогена. [При недостатке выработки инсулина формируется сахарный диабет].
А-клетки (ацидофильные) – составляют 20-25% всех клеток островка. Они располагаются по периферии инсулы. Они содержат гранулы, окрашивающиеся кислыми красителями. В электронном микроскопе гранулы имеют узкий ободок. Клетки также содержат развитый белоксинтезирующий аппарат и секретируют гормон глюкагон . Этот гормон является антагонистом инсулина (контринсулярный гормон), поскольку стимулирует распад гликогена в печени и способствует повышению содержания глюкозы в крови.
D -клетки составляют около 5% эндокринных клеток островка. Они располагаются по периферии инсулы. Содержат умеренно плотные гранулы без светлого ободка. В гранулах содержится гормон соматостатин , угнетающий функцию А, В-клеток островков и ациноцитов. Он же обладает митозингибирующим действием на различные клетки.
D 1-клетки содержат гранулы с узким ободком. Вырабатывают вазоин тестинальный полипептид , понижающий артериальное давление и стимулирующий выработку панкреатического сока. Количество этих клеток невелико.
 РР-клетки
(2-5%) располагаются по периферии
островков, иногда могут
встречаться и в составе экзокринной
части железы. Содержат гранулы различной
формы, плотности и величины. Клетки
вырабатывают панкреатический
полипептид
,
угнетающий внешнесекреторную активность
поджелудочной железы.
РР-клетки
(2-5%) располагаются по периферии
островков, иногда могут
встречаться и в составе экзокринной
части железы. Содержат гранулы различной
формы, плотности и величины. Клетки
вырабатывают панкреатический
полипептид
,
угнетающий внешнесекреторную активность
поджелудочной железы.
Поджелудочная железа считается одним из жизненно важных органов. Она не только активно участвует в процессе переваривания пищи, но и имеет эндокринные функции. Внутренняя секреция осуществляется благодаря специальным клеткам, расположенным в хвосте данного анатомического образования. Место, в котором вырабатываются гормоны поджелудочной железы, называется островок Лангерганса. Это анатомическое образование имеет огромное функциональное значение. Благодаря ему обеспечивается углеводный обмен.
Эндокринные клетки расположены по всему организму. Одним из мест их скопления является поджелудочная железа. Островки Лангерганса локализованы в хвостовой части органа. Они представляют собой клеточные скопления, вырабатывающие биологически активные вещества - гормоны. Значение островков Лангерганса огромное. Оно заключается в продукции нормального количества гормонов, необходимых для обменных процессов. Островки Лангерганса поджелудочной железы имеют следующие функции:
Благодаря нормальной работе островкового аппарата не развиваются такие состояния, как сахарный диабет и гипогликемия. Повреждение клеток возникает при острых и хронических воспалениях - панкреатитах.

Островок Лангерганса был открыт в 19 веке. Он представляет собой сосредоточение эндокринных элементов. У детей эти образования занимают около 6 % от общей площади органа. К взрослому возрасту эндокринная часть уменьшается и составляет всего 2 %. В паренхиме хвоста находится около миллиона островков Лангерганса. Они имеют собственное обильное кровоснабжение и иннервацию. Каждый островок состоит из долек, которые покрывает соединительная ткань. Кроме того, она находится и снаружи эндокринных образований. Клетки внутри островков расположены в виде мозаики. Активность эндокринных скоплений обеспечивают блуждающий и симпатический нервы. В центре дольки располагаются инсулярные клетки. Они снижают уровень глюкозы в крови. В периферической части долек находятся альфа- и дельта-клетки. Первые вырабатывают контринсулярный гормон - глюкагон. Вторые необходимы для регуляции эндокринной и экзокринной активности.

В островках Лангерганса вырабатывается несколько видов клеток. Все они участвуют в выделении биологически активных веществ - пептидов и гормонов. Большая часть островков Лангерганса представлена бета-клетками. Они располагаются в центре каждой дольки. Эти клетки очень важны, так как они вырабатывают инсулин.
Вторыми по значимости считаются альфа-клетки поджелудочной железы. Они занимают четверть площади островка. Альфа-клетки необходимы для продуцирования глюкагона. Этот гормон является антагонистом инсулина.
В периферийной части островков Лангерганса вырабатываются РР- и дельта-клетки. Количество первых составляет около 1/20 части. Функция данных образований - выработка полипептида поджелудочной железы. Дельта-клетки необходимы, чтобы продуцировать соматостатин. Данное вещество участвует в регуляции обмена углеводов.
Островковые клетки сложно поддаются регенеративным процессам. Поэтому при повреждении этих структур восстановить их функцию часто невозможно.

Несмотря на то, что островок Лангерганса имеет небольшой размер и занимает лишь малую часть поджелудочной железы, значение этого фрагмента велико. В нём происходит образование важнейших гормонов, участвующих в обменных процессах. Островки Лангерганса вырабатывают инсулин, глюкагон, соматостатин и панкреатический полипептид.
Первые 2 гормона необходимы для жизнедеятельности. Инсулин запускает процесс распада на глюкозы на более мелкие молекулярные соединения. В результате уровень сахара в крови снижается. Помимо этого инсулин участвует в обмене жиров. Благодаря действиям этого гормона в печени и мышечной ткани накапливается гликоген. Инсулин оказывает анаболический эффект на общий обмен веществ, то есть ускоряет все процессы.
Обратным действием обладает глюкагон. Этот гормон вырабатывается в меньшем количестве по сравнению с инсулином. Он участвует в глюконеогенезе. Сахар необходим в организме, так как является источником энергии.
Соматостатин регулирует выработку пищеварительных ферментов и гормонов. Под воздействием этого вещества снижается выработка глюкагона и инсулина. РР-клеток в островках Лангерганса очень мало, однако панкреатический полипептид для организма необходим. Он участвует в регуляции секреции пищеварительных желез (печени, желудка). При недостаточности гормональной активности развиваются тяжелые заболевания.

Нарушение деятельности островковых клеток может произойти по разным причинам. Зачастую недостаточность этих структур относится к врожденным аномалиям (генетическим патологиям). Приобретённое поражение островков Лангерганса развивается вследствие вирусных и бактериальных инфекций, хронической алкогольной интоксикации, неврологических заболеваний.
Недостаточность инсулина приводит к сахарному диабету 1-го типа. Эта болезнь возникает в детском и молодом возрасте. Повышение глюкозы в крови приводит к поражению сосудов и нервов. При дефиците других островковых клеток развивается гипогликемическое состояние, повышенная продукция пищеварительных соков. Усиленная выработка гормонов возникает при доброкачественных опухолях хвоста поджелудочной железы.
Методом лечения сахарного диабета является заместительная терапия инсулином. В последние годы разрабатываются альтернативные способы. К ним относятся внедрение искусственной поджелудочной железы и пересадка островковых клеток. На практике оказалось, что гормонпродуцирующие структуры приживаются в новом организме. При этом углеводный обмен может полностью восстановиться. Пересадка островков Лангерганса еще не получила широкого применения в практике.
В 19 веке молодой ученый из Германии обнаружил неоднородность тканей поджелудочной железы. Клетки, отличавшиеся от основной массы, располагались небольшими скоплениями, островками. Группы клеток в дальнейшем назвали именем патологоанатома — островки Лангерганса (ОЛ).
Их доля в общем объеме тканей составляет не более 1-2%, однако эта небольшая часть железы выполняет свою функцию, отличную от пищеварительной.
 Основная часть клеток поджелудочной железы (ПЖ) вырабатывает ферменты, способствующие пищеварению. Функция островных скоплений другая – они синтезируют гормоны, поэтому их относят к эндокринной системе.
Основная часть клеток поджелудочной железы (ПЖ) вырабатывает ферменты, способствующие пищеварению. Функция островных скоплений другая – они синтезируют гормоны, поэтому их относят к эндокринной системе.
Таким образом, поджелудочная железа является частью двух основных систем организма – пищеварительной и эндокринной. Островки являются микроорганами, вырабатывающими 5 видов гормонов.
Большая часть панкреатических групп расположены в хвостовой части поджелудочной железы, хотя хаотичные, мозаичные вкрапления захватывает всю экзокринную ткань.
ОЛ отвечают за регулирование углеводного обмена и поддерживают работу других эндокринных органов.
Каждый островок представляет собой самостоятельно функционирующий элемент. Вместе они составляют сложный архипелаг, который составлен из отдельных клеток и более крупных образований. Размеры их значительно различаются – от одной эндокринной клетки до зрелого, крупного островка (>100 мкм).
В панкреатических группах выстроена иерархия расположения клеток, их 5 видов, все выполняют свою роль. Каждый островок окружен соединительной тканью, имеет дольки, где находятся капилляры.
В центре расположены группы бета-клеток, по краям образований – альфа и дельта-клетки. Чем больше размер островка, тем больше в нем периферийных клеток.
Островки не имеют протоков, вырабатываемые гормоны выводятся по системе капилляров.

Разные группы клеток продуцируют свой вид гормона, регулируя пищеварение, липидный и углеводный обмен.
С течением жизни удельный вес эндокринной составляющей поджелудочной железы сокращается – от 6% в первые месяцы жизни до 1-2 % к 50-ти годам.
Гормональная роль поджелудочной железы велика.
Синтезированные в маленьких островках активные вещества током крови доставляются в органы и регулируют метаболизм углеводов:
Количество вырабатываемых гормонов зависит от полученной с пищей глюкозы и скорости ее окисления. При увеличении ее количества выработка инсулина увеличивается. Синтез запускается при концентрации 5,5 ммоль/л в плазме крови.
Спровоцировать выработку инсулина может не только прием пищи. У здорового человека максимальная концентрация отмечается в период сильных физических напряжений, стрессов.
Эндокринная часть ПЖ вырабатывает гормоны, которые оказывают решающее влияние на весь организм. Патологические изменения ОЛ способны нарушить работу всех органов.
Видео о задачах инсулина в организме человека:
Причиной поражения ОЛ могут быть генетическая предрасположенность, инфекции и отравления, воспалительные заболевания, иммунные проблемы.
В результате происходит прекращение или значительное снижение выработки гормонов разными клетками островков.
В результате этого могут развиться:
Основные принципы лечения сахарного диабета 1 типа заключаются во введение в организм инсулина, выработка которого нарушена или снижена. Применяют два вида инсулинов – быстрый и длительного действия. Последний вид имитирует выработку гормона ПЖ.
 СД 2 типа требует строгого соблюдения диеты, умеренных физических упражнений и приема препаратов, способствующих сжиганию сахара.
СД 2 типа требует строгого соблюдения диеты, умеренных физических упражнений и приема препаратов, способствующих сжиганию сахара.
Во всем мире наблюдается рост заболеваемости диабетом, его уже называют чумой 21 века. Поэтому медицинские исследовательские центры ищут способы борьбы с заболеваниями островков Лангерганса.
Процессы в поджелудочной железе развиваются быстро и приводят к отмиранию островков, которые должны синтезировать гормоны.
В последние годы стало известно:
Это позволяет пациентам отказаться от постоянного приема лекарственных средств, строгой диеты и вернуть нормальный образ жизни. Проблемой остается иммунная система, которая может отторгнуть подсаженные клетки.
Еще одним возможным способом лечения рассматривается пересадка от донора части островковой ткани. Этот способ заменяет установку искусственной ПЖ или ее полную пересадку от донора. При этом удается остановить прогрессирование заболевания и нормализовать глюкозу в крови.
Проведены успешные операции, после которых у больных с диабетом 1 типа отпала необходимость во введении инсулина. Орган восстановил популяцию бета-клеток, синтез собственного инсулина возобновился. После операции была проведена иммуносупрессивная терапия, чтобы не допустить отторжения.
Видео-материал о функциях глюкозы и заболевании диабетом:
Медицинские институты работают над изучением возможности пересадки поджелудочной железы от свиньи. Первые средства для лечения сахарного диабета как раз использовали части поджелудочной железы свиней.
Ученые сходятся на том, что необходимы исследования особенностей строения и работы островков Лангерганса из-за большого количества важных функций, которые выполняют синтезируемые в них гормоны.
Постоянный прием искусственных гормонов не помогает победить болезнь и ухудшает качество жизни пациента. Поражение этой маленькой части ПЖ вызывает глубокие нарушения работы всего организма, поэтому исследования продолжаются.
Человеческий организм – разумный и достаточно сбалансированный механизм.

Среди всех известных науке инфекционных заболеваний, инфекционному мононуклеозу отводится особое место...

О заболевании, которое официальная медицина называет «стенокардией», миру известно уже достаточно давно.

Свинкой (научное название – эпидемический паротит) называют инфекционное заболевание...

Печеночная колика является типичным проявлением желчнокаменной болезни.

Отек головного мозга – это последствия чрезмерных нагрузок организма.

В мире не существует людей, которые ни разу не болели ОРВИ (острые респираторные вирусные заболевания)...

Здоровый организм человека способен усвоить столько солей, получаемых с водой и едой...
Бурсит коленного сустава является широко распространённым заболеванием среди спортсменов...
Поджелудочная железа выполняет разные функции. Одна из них эндокринная, то есть этот орган вырабатывает гормоны. Такая функция поджелудочной железы обеспечивается специальными клетками, которые предназначены именно для этого.
Эндокринная функция поджелудочной железы обеспечивается работой скопления клеток, имеющих эпителиальное происхождение. Эти скопления называют островками Лангерганса, они составляют 1-2% от всего органа. Количество таких островков в железе у взрослого человека от двухсот тысяч до полутора миллионов. Клетки островков Лангерганса бывают трех видов и продуцируют различные гормоны.
Типы клеток и гормоны, которые они продуцируют:
Альфа-клетки поджелудочной железы вырабатывают глюкагон. Этот гормон отвечает за многие процессы:
Данный гормон вырабатывают дельта-клетки островков Лангерганса поджелудочной железы. Его биологическая роль - подавлять секрецию соматотропного гормона, глюкагона, инсулина и некоторых других гормонов, а также электролитов, панкреатических ферментов, желудочного сока. Кроме того, под воздействием этого гормона замедляются кровоток внутренних органов, перистальтика кишечника, а также возбудимость нервных окончаний. Таким образом, за счет увеличения или уменьшения количества соматостатина регулируется необходимый уровень других гормонов и работа некоторых внутренних органов.
О гормоне инсулин, который вырабатывается бета-клетками поджелудочной железы, знают очень многие. Он нужен нам для расщепления глюкозы и выработки энергии в организме. Продукция этого гормона обеспечивается за счет взаимодействия глюкозы с различными рецепторами, в реакции участвуют также некоторые аминокислоты.
Основное влияние инсулин в нашем организме оказывает на углеводный обмен. Под его воздействием увеличивается транспорт глюкозы в клетки тканей, которые являются инсулинозависимыми. Это ткани печени, мышц, а также жировая ткань. Непосредственного действия на нервные ткани, почки инсулин не оказывает, однако нарушение баланса сахара в крови при недостатке или избытке инсулина может оказать разрушительное действие на все органы.
Кроме регуляции углеводного обмена, инсулин участвует и в других видах обмена. К примеру, он стимулирует транспорт аминокислот через клеточные мембраны, участвует в синтезе белка и тормозит его распад. При регуляции жирового обмена за счет количества инсулина происходит включение жирных кислот в жировую ткань, корректируется синтез липидов и липолиз.
Инсулин способен связываться с особыми рецепторами клеточной мембраны. После их соединения сигнал передается в систему цАМФ через фермент оболочки клетки аденилатциклазу. Эта система регулирует синтез белка и отвечает за утилизацию глюкозы.
Все гормоны важны для поддержания функций организма. Однако в энергетическом балансе основная роль принадлежит инсулину и глюкагону.
Именно эти гормоны помогают поддерживать энергию на определенном уровне. За счет увеличения и уменьшения продукции то одного из этих гормонов, то другого организм обеспечивает нормальный уровень сахара. Если происходит нарушение способности клеток островков Лангерганса продуцировать эти гормоны, или существенно уменьшается их количество, в организме могут быть серьезные сбои и развиваться заболевания.
moyaschitovidka.ru
Эндокринная часть поджелудочной железы представлена островками секреторных клеток (островками Лангерганса), расположенными между экзокринными ацинусами (см. Атл.). Больше островков в хвостовой части железы. Общее их число - 1-2 млн и более, но все же их объем не превышает 3% объема железы. Островки бывают овальной, лентовидной или звездчатой формы. С возрастом количество островков уменьшается.
Обновление клеток островкового аппарата происходит за счет медленного их деления. При избытке углеводов в рационе человека и животных клетки, вырабатывающие инсулин, испытывают повышенную нагрузку. В результате такой гиперфункции начинается их гибель. Вследствие этого развивается заболевание, названное сахарным диабетом. Инсулин и глюкагон участвуют во всех видах обмена веществ.
Выделяют четыре основных типа эндокринных клеток поджелудочной железы, каждый из которых синтезирует один определенный гормон:

Дифференцировка клеток, синтезирующих инсулин и глюкагон, происходит в течение 3 месяцев внутриутробного развития, на 12 неделе проявляется их секреторная активность, а к концу 5 месяца островки Лангерганса приобретают характерное для взрослых строение.
Инсулин совместно с гормоном роста регулирует ростовые процессы: его концентрация повышается в периоды интенсивного роста и после рождения.
doctor-v.ru
В эндокринной части паренхимы поджелудочной железы располагаются островки Лангерганса. Их основными структурными единицами являются секреторные (α, β, Δ, F и другие) клетки.
А-клетки (α-клетки) островков продуцируют глюкагон. Он увеличивает гликогенолиз в печени, снижает в ней утилизацию глюкозы, а также повышает глюконеогенез и образование кетоновых тел. Результатом этих воздействий является увеличение концентрации глюкозы в крови. Вне печени глюкагон повышает липолиз и снижает синтез белков.
На -клетках имеются рецепторы, которые при уменьшении уровня глюкозы во внеклеточной среде усиливают секрецию глюкагона. Секретин угнетает продукцию глюкагона, а другие желудочно-кишечные гормоны стимулируют ее.
B-клетки (-клетки) синтезируют и накапливают инсулин. Этот гормон увеличивает проницаемость клеточных мембран для глюкозы и аминокислот, а также способствует превращению глюкозы в гликоген, аминокислот в белки, а жирных кислот в триглицериды.
Синтезирующие инсулин клетки способны реагировать на изменения содержания в крови и просвете ЖКТ калоригенных молекул (глюкозы, аминокислот и жирных кислот). Из аминокислот наиболее выражена стимуляция секреции инсулина аргинином и лизином.
Поражение островков Лангерганса приводит к гибели животного из-за нехватки в организме инсулина. Только этот гормон снижает содержание глюкозы в крови.
Д-клетки (Δ-клетки) островков синтезируют панкреатический соматостатин. В поджелудочной железе он оказывает тормозящее паракринное влияние на секрецию гормонов островками Лангерганса (преобладает влияние на -клетки), а внешнесекреторным аппаратом - бикарбонатов и ферментов.
Эндокринное влияние панкреатического соматостатина проявляется торможением секреторной активности в ЖКТ, аденогипофизе, паращитовидной железе и почках.
Наряду с секрецией, панкреатический соматостатин снижает сократительную активность желчного пузыря и желчных протоков, а на всем протяжении ЖКТ - уменьшает кровообращение, моторику и всасывание.
Активность Д-клеток возрастает при высоком содержании в просвете пищеварительного тракта аминокислот (особенно лейцина и аргинина) и глюкозы, а также при увеличении концентрации в крови ХКП, гастрина, желудочного ингибирующего полипептида (ЖИП) и секретина. В то же время, норадреналин угнетает высвобождение соматостатина.
Панкреатический полипептид синтезируется F-клетками (или РР-клетками) островков. Он уменьшает объем панкреатического секрета и концентрацию в нем трипсиногена, а также тормозит выведение желчи, но стимулирует базальную секрецию желудочного сока.
Выработка панкреатического полипептида стимулируется парасимпатической нервной системой, гастрином, секретином и ХКП, а также при голодании, приеме богатого белками корма, гипогликемии и физической нагрузке.
Интенсивность выработки гормонов поджелудочной железы контролируется вегетативной нервной системой (парасимпатические нервы вызывают гипогликемию, а симпатические - гипергликемию). Однако основными факторами регуляции секреторной активности клеток в островках Лангерганса, являются концентрации питательных веществ в крови и просвете ЖКТ. Благодаря этому, своевременные реакции клеток островкового аппарата обеспечивают поддержание постоянного уровня питательных веществ в крови между приемами корма.
ЭНДОКРИННАЯ ФУНКЦИЯ ПОЛОВЫХ ЖЕЛЕЗ
После наступления половой зрелости основными источниками половых гормонов в организме животных становятся постоянные половые железы (у самцов - семенники, а у самок - яичники). У самок периодически могут появляться и временные эндокринные железы (например, плацента во время беременности).
Половые гормоны делят на мужские (андрогены) и женские (эстрогены).
Андрогены (тестостерон, андростендион, андростерон и др.) специфически стимулируют рост, развитие и функционирование органов размножения самцов, а с наступлением половой зрелости - образование и созревание мужских половых клеток.
Еще до рождения в организме плода формируются вторичные половые признаки. Это в значительной степени регулируется образующимися в семенниках андрогенами (секретируются клетками Лейдига) и фактором, секретируемым клетками Сертоли (находятся в стенке семенного канальца). Тестостерон обеспечивает дифференцировку наружных половых органов по мужскому типу, а секрет клеток Сертоли предотвращает образование матки и маточных труб.
В период полового созревания андрогены ускоряют инволюцию тимуса, а в других тканях стимулируют накопление питательных веществ, синтез белка, развитие мышечной и костной ткани, повышают физическую работоспособность и сопротивляемость организма неблагоприятным воздействиям.
Андрогены влияют на ЦНС (например, вызывают проявления полового инстинкта). Поэтому удаление половых желез (кастрация) у самцов делает их спокойными и может привести к нужным для хозяйственной деятельности изменениям. Например, кастрированные животные быстрее откармливаются, мясо их вкуснее и нежнее.
До рождения, секреция андрогенов обеспечивается совместным действием на плод ЛГ самки и хорионического гонадотропина (ХГ). После рождения, развитие семенных канальцев, спермиев и сопровождающую эти процессы выработку БАВ клетками Сертоли стимулирует собственный гонадотропин самца - ФСГ, а ЛГ вызывает секрецию тестостерона клетками Лейдига. Старение сопровождается угасанием активности половых желез, но продолжается выработка половых гормонов надпочечником.
К видовым особенностям клеток Сертоли семенников жеребца, быка и кабана относится их способность кроме тестостерона вырабатывать эстрогены, которые регулируют обмен веществ в половых клетках.
Яичники в организме половозрелой самки в соответствии со стадиями полового цикла вырабатывают эстрогены и гестагены. Основным источником эстрогенов (эстрона, эстрадиола и эстриола) являются фолликулы, а гестагенов - желтое тело.
У неполовозрелой самки эстрогены надпочечников стимулируют развитие репродуктивной системы (яйцеводов, матки и влагалища) и вторичных половых признаков (определенного телосложения, молочных желез и т.д.). После наступления половой зрелости, концентрация в крови женских половых гормонов значительно повышается за счет их интенсивной выработки яичниками. Возникающие при этом уровни эстрогенов стимулируют созревание половых клеток, синтез белков и образование мышечной ткани в большинстве внутренних органов самки, а также повышают сопротивляемость ее организма к вредным воздействиям и вызывают связанные с половыми циклами изменения в органах животного.
Высокие концентрации эстрогена вызывают рост, расширение просвета и усиление сократительной активности яйцеводов. В матке они повышают кровенаполнение, стимулируют размножение клеток эндометрия и развитие маточных желез, а также изменяют чувствительность миометрия к окситоцину.
У самок многих видов животных эстрогены вызывают ороговение клеток влагалищного эпителия перед течкой. Поэтому качество гормональной подготовки самки к спариванию и овуляции выявляют по цитологическим анализам вагинального мазка.
Эстрогены также способствуют формированию состояния «охоты» и соответствующих половых рефлексов в наиболее благоприятную для оплодотворения стадию полового цикла.
После овуляции, на месте бывшего фолликула образуется желтое тело. Вырабатываемые им гормоны (гестагены) влияют на матку, молочные железы и ЦНС. Они вместе с эстрогенами регулируют процессы зачатия, имплантации оплодотворенной яйцеклетки, вынашивания беременности, родов и лактации. Основным представителем гестагенов является прогестерон. Он стимулирует секреторную активность маточных желез и делает эндометрий способным реагировать на механические и химические воздействия разрастаниями, которые необходимы для имплантации оплодотворенной яйцеклетки и образования плаценты. Прогестерон также снижает чувствительность матки к окситоцину и расслабляет ее. Поэтому преждевременное снижение концентрации гестагенов в крови беременных самок вызывает роды до полного созревания плода.
Если беременность не наступила, то желтое тело подвергается инволюции (продукция гестагенов прекращается) и начинается новый овариальный цикл. Умеренные количества прогестерона в синергизме с гонадотропинами стимулируют овуляцию, а большие - тормозят секрецию гонадотропинов и овуляция не происходит. Небольшие количества прогестерона также необходимы для обеспечения течки и готовности к спариванию. Кроме этого, прогестерон участвует в формировании доминанты беременности (гестационной доминанты), направленной на обеспечение развития будущего потомства.
После воздействия эстрогенов, прогестерон способствует развитию железистой ткани в молочной железе, что приводит к формированию в ней секреторных долек и альвеол.
Наряду со стероидными гормонами желтое тело, эндометрий и плацента, преимущественно перед родами, продуцируют гормон релаксин. Его выработка стимулируется высокими концентрациями ЛГ и вызывает повышение эластичности лонного сочленения, расслабление связки тазовых костей, а непосредственно перед родами повышает чувствительность миометрия к окситоцину и вызывает расширению маточного зева.
Плацента возникает в несколько этапов. Сначала, в ходе дробления оплодотворенной яйцеклетки образуется трофобласт. После присоединения к нему внезародышевых кровеносных сосудов трофобласт превращается в хорион, который после плотного соединения с маткой становится сформировавшейся плацентой.
У млекопитающих плацента обеспечивает прикрепление, иммунологическую защиту и питание плода, выведение продуктов обмена, а также выработку гормонов (эндокринная функция), необходимых для нормального течения беременности.
Уже на ранних сроках беременности в местах прикрепления ворсинок хориона к матке вырабатывается хорионический гонадотропин. Его появление ускоряет развитие зародыша и предотвращает инволюцию желтого тела. Благодаря этому желтое тело поддерживает высокий уровень прогестерона в крови до тех пор, пока плацента сама не начнёт синтезировать его в необходимом количестве.
Вырабатываемые в организме беременных самок негипофизарные гонадотропины имеют видовые особенности, но могут влиять на репродуктивные функции и у других видов животных. Например, введение гонадотропина сыворотки крови жеребых кобыл (ГСЖК) вызывает у многих млекопитающих выделение прогестерона. Это сопровождается удлинением полового цикла и задерживает приход охоты. У коров и овец ГСЖК также вызывает одновременный выход нескольких зрелых яйцеклеток, что используется при трансплантации эмбрионов.
Плацентарные эстрогены вырабатываются плацентой большинства млекопитающих (у приматов - эстрон, эстрадиол и эстриол, а у лошади - эквилин и эквиленин) преимущественно во второй половине беременности из дегидроэпиандростерона образующегося в надпочечниках плода.
Плацентарный прогестерон у ряда млекопитающих (приматы, хищники, грызуны) секретируются в количествах достаточных для нормального вынашивания плода даже после удаления желтых тел.
Плацентарный лактотропин (плацентарный лактогенный гормон, плацентарный пролактин, хорионический соматомаммотропин) поддерживает рост плода, а у самки увеличивает синтез белка в клетках и концентрацию СЖК в крови, стимулирует рост секреторных отделов молочных желёз и их подготовку к лактации, а также задерживает в организме ионы кальция, снижает мочевую экскрецию фосфора и калия.
По мере увеличения сроков беременности в крови самок растет уровень плацентарного кортиколиберина, который увеличивает чувствительность миометрия к окситоцину. Данный либерин практически не влияет на секрецию АКТГ. Это связано с тем, что во время беременности в крови растет содержание белка, который быстро нейтрализует кортиколиберин и он не успевает подействовать на аденогипофиз.
Тимус (зобная или вилочковая железа) имеется у всех позвоночных животных. У большинства млекопитающих он состоит из двух соединенных друг с другом долей, расположенных в верхней части грудной клетки сразу за грудиной. Однако, у сумчатых животных эти доли тимуса обычно остаются отдельными органами. У пресмыкающихся и птиц железа обычно имеет вид цепочек, расположенных по обе стороны шеи.
Наибольших размеров по отношению к массе тела тимус большинства млекопитающих достигает к моменту рождения. Затем он медленно растет и в период полового созревания достигает максимальной массы. У морских свинок (и некоторых других видов животных) крупный тимус сохраняется на протяжении всей жизни, но у большинства высокоразвитых животных после полового созревания железа постепенно уменьшается (физиологическая инволюция), но полной атрофии ее не происходит.
В тимусе эпителиальные клетки продуцируют тимические гормоны влияющие эндокринным и паракринным путем на гемопоэз, а также дифференцировку и активность Т-клеток.
В тимусе на предшественники Т-лимфоцитов последовательно действуют тимопоэтин и тимозины. Они делают дифференцирующиеся в тимусе клетки чувствительными к активированному кальцием тимулину (или тимическому сывороточному фактору - ТСФ).
П р и м е ч а н и е: Возрастное снижение содержания ионов кальция в организме является причиной падения активности тимулина у старых животных.
Секреторная активность тимуса тесно связана с деятельностью гипоталамуса и других эндокринных желез (гипофиза, эпифиза, надпочечников, щитовидной железы и гонад). Гипоталамический соматостатин, удаление надпочечников и щитовидной железы снижают выработку тимических гормонов, а эпифиз и кастрация усиливают гормонопоэз в тимусе. Кортикостероиды регулируют распределение тимических гормонов между тимусом, селезенкой и лимфоузлами, а тимэктомия приводит к гипертрофии коры надпочечников.
Перечисленные примеры свидетельствуют о том, что вилочковая железа обеспечивает интеграцию нейро-эндокринной и иммунной систем в целостном макроорганизме.
Эпифиз (шишковидная железа) расположена у позвоночных под кожей головы или в глубине мозга. Основными клетками эпифиза у млекопитающих являются пинеалоциты, а у более примитивных животных здесь имеются и фоторецепторы. Поэтому, наряду с эндокринной функцией эпифиз может обеспечивать ощущение степени освещенности объектов. Это позволяет глубоководным рыбам осуществлять вертикальную миграцию в зависимости от смены дня и ночи, а миногам и пресмыкающимся - оберегать себя от опасности сверху. У некоторых перелетных птиц эпифиз, вероятно, выполняет функцию навигационных приборов при перелетах.
Эпифиз земноводных уже способен вырабатывать гормон мелатонин, который уменьшение количество пигмента в клетках кожи.
Пинеалоциты непрерывно синтезируют гормон серотонин, который в темное время суток и при низкой активности симпатической нервной системы (у птиц и млекопитающих) превращается в мелатонин. Поэтому продолжительность дня и ночи, влияют на содержание этих гормонов в эпифизе. Возникающие при этом ритмические изменения их концентрации в шишковидной железе определяют у животных суточный (циркадианный) биологический ритм (например, периодичность сна и колебания температуры тела), а также влияет на формирование таких сезонных реакций как зимняя спячка, миграция, линька и размножение.
Увеличение содержания мелатонина в эпифизе оказывает снотворный, анальгезирующий и седативный эффекты, а также тормозит половое созревание молодняка. Поэтому после удаления эпифиза у цыплят быстрее наступает половое созревание, у самцов млекопитающих - гипертрофируются семенники и усиливается созревание спермиев, а у самок - удлиняется период жизни желтых тел и увеличивается матка.
Мелатонин снижает секрецию ЛГ, ФСГ, пролактина и окситоцина. Поэтому низкий уровень мелатонина в светлое время суток способствует усилению молокообразования и высокой половой активности животных в те времена года, когда ночи наиболее короткие (весной и летом). Мелатонин также нейтрализует повреждающее действие стрессоров и является естественным антиоксидантом.
У млекопитающих серотонин и мелатонин выполняют свои функции в основном в эпифизе, а дистантными гормонами железы, вероятно, являются полипептиды. Значительная их часть наряду с кровью, секретируется в спинномозговую жидкость и через нее поступает в различные отделы ЦНС. Это оказывает преимущественно тормозное влияние на поведение животного и другие функции мозга.
В эпифизе уже обнаружено около 40 секретирующихся в кровь и спиномозговую жидкость биологически активных пептидов. Из них наиболее изучены антигипоталамические факторы и адреногломерулотропин.
Антигипоталамические факторы обеспечивают связь эпифиза с гипоталамо-гипофизарной системой. К ним, например, относятся аргинин-вазотоцин (регулирует секрецию пролактина) иантигонадотропин(ослабляет секрецию ЛГ).
Адреногломерулотропин стимулируя выработку альдостерона надпочечником, влияет на водно-солевой обмен.
Таким образом, основной функцией эпифиза является регуляция и координация биоритмов. Посредством контроля деятельности нервной и эндокринной систем животного, шишковидная железа обеспечивает опережающую реакцию его систем на смену времени суток и сезона.
studfiles.net
Поджелудочная (панкреатическая) железа относится к органам с двойной секрецией. Внешнесекреторный аппарат железы вырабатывает составные части панкреатического сока, экскретируемо-го в двенадцатиперстную кишку. Около 1,5-2% массы железы приходится на эндокринную ткань (островки Лангерганса) - группы скоплений специальных паренхиматозных клеток. Кровоснабжение поджелудочной железы осуществляется поджелудочно-двенадцатиперстной артерией и ветвями селезеночной артерии, причем кровоснабжение островков Лангерганса существенно обильнее, чем других частей органа. Вены поджелудочной железы впадают в воротную вену через селезеночную или верхнюю брыжеечную вену. Иннервируется железа ветвями блуждающего и симпатического нервов.
В островках Лангерганса имеется несколько видов клеток: β-клетки, расположены ближе к центру островков и составляют до 60-70 % всех клеток; δ-клетки (2-8 %) - предшественники других клеток островков и α-клетки (около 25 %), находятся ближе к периферии островков. Протоплазма α- и β-клеток содержит гранулы, а δ-клетки негранулированы. α-Клетки неаргирофильны и являются местом образования глюкагона; β-клетки образуют инсулин, δ-клетки - соматотропин. РР-клетки, также имеющиеся в железе, расположены по периферии островков и в паренхиме возле протоков малого и среднего диаметра. Они секретируют панкреатический полипептид. В островках выявлено некоторое количество клеток - продуцентов вазоактивного интерстициального пептида (ВИП) и гастроинтерстициального пептида (ГИП).
Инсулин - низкомолекулярный белок с молекулярной массой около 6000 Д. В его состав входит 16 аминокислот и 51 аминокислотный остаток. В настоящее время синтезирован искусственным путем. Он образуется из проинсулина под влиянием протеаз; его активность составляет около 5 % активности инсулина. Считается, что биологический эффект инсулина связан с его способностью соединяться со специфическими рецепторами цитоплазматических мембран клеток, после чего передается сигнал на систему цАМФ через фермент аденилатциклазу оболочки клетки цАМФ, который регулирует синтез белка и утилизацию глюкозы при участии Са++ и Мg++.
С кровью инсулин поступает в печень, где около половины его инактивируется под воздействием инсулиназы, а остальная часть связывается с белками, частично оставаясь свободной.
Из печени инсулин поступает в кровь в свободном и связанном с белками состоянии. Это соотношение регулируется уровнем гликемии. При понижении сахара в крови преобладает белковосвязанная фракция, а при гипергликемии - свободный инсулин, который действует на инсулиночувствительные субстанции, а связанная фракция - только на жировую ткань, в которой имеются пептидазы, освобождающие инсулин из связанного состояния. Период полураспада инсулина - около 30 мин. Инсулин кроме печени инактивируется в жировой ткани, мышцах, почках, плаценте.
Основным биостимулятором синтеза инсулина является глюкоза, под влиянием которой в поджелудочной железе синтез инсулина повышается, а с уменьшением ее - снижается.
Стимуляторами освобождения и секреции инсулина являются также СТГ, АКТЕ, глюкокортикоиды, глюкагон, секретин, аргинин, лейцин, гастрин, бомбезин, панкреозимин, желудочный ингибитор - полипептид, нейротензин, β-адреностимуляторы, сульфаниламиды, соматостатин.
Соматостатин - 14-членный пептид, обнаружен в гипоталамусе, образуется также в δ-клетках островков Лангерганса, клетках щитовидной железы, желудка и лимфоидных органов. Он подавляет секрецию ТТГ, СТГ, АКТГ, гастрина, секретина, мотилина, ренина, вазоактивного желудочного пептида (ВЖП), панкреатических ферментов, желудочного сока; снижает перистальтику кишечника, сократимость мочевого пузыря, абсорбцию ксилозы. Под его влиянием уменьшается освобождение ацетилхолина из нервных окончаний и электровозбудимость нервов. Является ингибитором секреции инсулина и глюкагона. Парасимпатическая стимуляция увеличивает секрецию инсулина, а симпатическая - уменьшает. Важную роль в секреции инсулина играют холинэргитические волокна блуждающего нерва.
Инсулинстимулирует перенос Сахаров через мембрану клеток жировой, мышечной, почечной тканей; усиливает фосфорилирование, окисление и превращение глюкозы в гликоген и жиры; способствует превращению жирных кислот в триглицириды жировой ткани; стимулирует синтез липидов; ингибирует липолиз и активность глюкозо-6-фосфатазы; стимулирует образование макроэргических связей, транспорт аминокислот через цитоплазматические мембраны; ослабляет глюкогенолиз из белка; способствует его синтезу из аминокислот. Все ткани, кроме нервной, сетчатки, почечной и эритроцитов, чувствительны к инсулину.
Глюкагонявляется антагонистом инсулина. Это полипептид, состоящий из 29 аминокислотных остатков с молекулярной массой 3485 Д. Он усиливает распад гликогена в печени и тормозит его синтез; усиливает липолиз, гликонеогенез, биосинтез глюкозы из аминокислот; способствует снижению кальциемии и фосфатемии, выходу калия из печени, отчего наступает значительная, но скоротечная гиперкалиемия, сменяющаяся затем гипокалиемией, которая обусловлена гиперкалийурией и усилением депонирования калия клетками.
Секреция глюкагона снижается при гипергликемии, повышении в крови свободных жирных кислот и под влиянием соматостатина.
Глюкагон тормозит агрегацию тромбоцитов, способствует увеличению минутного объема кровотока. Под его влиянием увеличивается образование СТГ, инсулина, катехоламинов, кальцитонинов, выделение воды и электролитов с мочой, а секреция панк-реозимина, гастрина, панкреатических ферментов снижается.
Кроме панкреатического глюкагона известен также кишечный глкжагон, секретируемый α-клетками слизистой оболочки желудка и кишечника. Он усиливает липолиз, гликогенолиз, стимулирует секрецию инсулина. Секреция кишечного глюкагона повышается при поступлении в кишечник пищи и соединений кальция.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
И УГЛЕВОДНЫЙ ОБМЕН
Углеводы являются основным энергетическим материалом, реализующимся при распаде глюкозы в цикле Кребса (аэробном цикле трикарбоновых кислот) на Н2О и СО2. Образование гликогена из моно- и дисахаридов, гексоз и пентоз происходит под влиянием инсулина, а основное количество углеводов у жвачных расщепляется в преджелудках под воздействием микрофлоры до ЛЖК, а у моногастричных - в тонком кишечнике под влиянием ферментов поджелудочной железы (мальтазы, амилазы, лактазы) до моносахаридов. Более 85 % моносахаридов переходят в глюкозу уже в тонком кишечнике и около 15 % - в печени. В процессах фосфорилирования глюкоза является активным звеном окисления, синтеза гликогена и жира. На первом этапе фосфорилирования образуется гексозомонофосфат:
глюкоза + АТФ -> гексакиназа -> гексозомонофосфат + АДФ.
Особенностью этого превращения является то, что к молекуле глюкозы присоединяется не простая (неорганическая), а обогащенная энергией фосфорная кислота (макроэргическая связь), что делает глюкозу биологически активной, причем активатором гексокиназы в этом процессе является инсулин. Проникая через стенку кишечника и под влиянием фосфатазы дефосфорилируясь, глюкоза поступает в портальное кровообращение, теряя физиологическую активность. В печени она вторично фосфорилируется, образуя глюкозо-6-фосфат (Г-6-Ф), становясь снова физиологически активной под действием инсулина, и образует гликоген. Значение этого цикла в том, что он является единственным источником рибозо-5-фосфата, используемого в синтезе РНК. При окислении глюкозы в пентозном цикле образуется основная часть восстановленного NАДН - никотинамидадениндинуклеотида, необходимого для синтеза жирных кислот. В анаэробном цикле окисляется около 25 % Г-6-Ф, а около 55 % под влиянием глкжозо-6-фосфатазы, освобождаясь от фосфорной кислоты, из печени переходит в общий проток. 9 % из 55 (принятых за 100 %) этой глюкозы превращается в гликоген мышечной ткани, а около 30 % - в жир. Основная часть глюкозы (около 60 %) окисляется в тканях, обеспечивая энергетический баланс организма в анаэробном (с образованием молочной кислоты) и аэробном (с образованием Н2О и СО2) циклах. Молочная кислота в печени и мышцах может ресинтезироваться в гликоген, а образовавшаяся в аэробном гликолизе пировиноградная кислота декарбоксилируется с образованием ацетилкоэнзима А (ацетил-КоА), который необходим в дальнейшем синтезе жирных кислот, кетоновых (ацетоновых) тел, холестерина. В цикле ди- и трикарбоновых кислот в легких, почках, мышцах и частично в печени ацетил-КоА окисляется до Н2О и СО2, а катализатором этого процесса является инсулин. Аэробный гликолиз является наиболее эффективным - в его процессе образуется 36 молекул аденозинтрифосфорной кислоты (АТФ), тогда как в анаэробном только две молекулы АТФ.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
И ЛИПИДНЫЙ ОБМЕН
Основной резерв энергии организма - жиры. Из жировых депо жиры в виде свободных неэстерифицированных жирных кислот (НЭЖК) поступают в кровь, а затем в печень, где диализируются и используются тканями как энергетический материал. НЭЖК доставляют около 50 % тепловой энергии основного обмена.
Триглицериды жировых депо, поступая в кровь, образуют комплексы с α- и β-глобулинами и затем выходят из них в виде α- и β-липопротеидов. В норме жир в печени не задерживается, а откладывается в жировых депо. Этот процесс активируется гепарином, продуцируемым тучными клетками. Нормальными промежуточными продуктами обмена НЭЖК являются ацетоновые (кетоновые) тела, содержание которых в крови здоровых животных составляет в среднем 2-7 мг%. Кетоновые тела образуются в основном в печени. Усиленный кетоногенез (при недостаточности аэробного цикла, энергетическом голодании) - причина ацетонемии, кетоза, являющихся причиной дистрофии внутренних органов (миокарда, почек, печени), яловости, ацетонурии, ацетонолактии, «голодных» кетозов овец и свиней.
Непосредственно участвуют в обмене жиров фосфолипиды, способствующие окислению жира через стадию лецитина. Они же повышают устойчивость холестерина в крови, что препятствует его отложению в стенках сосудов.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
И БЕЛКОВЫЙ ОБМЕН
Более половины белков сыворотки крови (6-8 г%) составляют альбумины. Остальная часть их представлена α1-, α2-, β- и γ-глобулинами.
Альбумины синтезируются в паренхиматозных клетках печени, а глобулины - в ретинулоэндотелиальной системе (РЭС). Все питательные вещества в процессе обмена между кровью и клетками тканей проходят через основное вещество соединительной ткани, важнейшими элементами которой являются коллагеновые и эластические волокна белковой природы. Из этого следует, что любой фактор или состояние, влияющие на обмен белка, оказывают воздействие и на них.
Высокомолекулярные линейные полиэлектролиты соединительной ткани называются кислыми мукополисахаридами, а в соединении с белком - мукопротеидами (мукополисахаридными комплексами). В крови имеются также гликопротеиды - белки с содержанием около 4 % избытка глюкозамина.
Повышение белкового синтеза происходит под влиянием инсулина вследствие усиления переноса аминокислот в цитоплазму, активации ферментов пептидного цикла и усиления утилизации глюкозы (источника энергии макроэргических связей). Наряду с инсулином синтез белка стимулирует соматотропный гормон гипофиза (СТГ). Наоборот, АКТЕ, ТТГ, глюкокортиноиды, гормоны щитовидной железы стимулируют диализ белка до аминокислот.
САХАРНЫЙ ДИАБЕТ
Сахарный диабет представляет собой синдром хронической гипергликемии вследствие генетических и экзогенных факторов на почве абсолютного или относительного дефицита инсулина, сопровождающийся нарушением промежуточного обмена, особенно углеводного. Принято выделять три пути развития инсулинзави-симого сахарного диабета: 1) предрасположенность к аутоиммунному нарушению островков Лангерганса; 2) повышенная чувствительность β-клеток к вирусам и 3) ослабление противовирусного иммунитета. Чаще возникает в критические периоды - максимального роста и продуктивности, гормональной, иммунологической и других видов перестройки.
Сахарный диабет может возникать вторично - при панкреатитах, кистах, опухолях поджелудочной железы, гемохроматозах, особенно при эндокринных нарушениях других желез внутренней секреции, от ятрогенных причин, длительного применения диуретиков (особенно диазидов, кортикостероидов), при нарушениях кормления (длительное кормление турнепсом, брюквой, репой, капустой). Он чаще бывает вследствие относительной внепанкреатической инсулиновой недостаточности, чем абсолютной (панкреатической).
Патогенез инсулинзависимого сахарного диабета связан с деструкцией β-клеток, что приводит к абсолютному недостатку инсулина - «вирусному» или аутоиммунному. Повреждение более 90 % клеток поджелудочной железы приводит к развитию клинических симптомов диабета.
При дефиците инсулина понижается проницаемость для глюкозы цитоплазматических мембран в мышечной и жировой тканях, снижается ее фосфорилирование и окисление глюкозы, переход в спирт, усиливается гликонеогенез из белка и выделение углеводов из печени в кровь. Это приводит к неполной утилизации углеводов тканями - гипергликемии. В крови повышается содержание молочной кислоты - продукта анаэробного гликолиза. Возникает глюкозурия, полидипсия, ацетонемия, гипергликемия, что приводит к повышению осмотического давления крови и нарушению функций ЦНС. Нарушается липидный обмен (увеличение содержания в крови НЭЖК). Печень подвергается жировой дистрофии. Возрастает холестеринемия. Снижение концентрации фосфолипидов, гиперхолестеринемия, повышение содержания β-липопротеидов при диабете предрасполагают к ангиопатиям, атеросклерозу. Липоидозу способствует уменьшение расщепления триглицеридов в стенке сосудов, нарушается синтез, усиливается распад белков. Содержание альбуминов снижается, α1-, β - и γ-глобулинов повышается, что связано как с недостатком инсулина, так и с недостаточностью гипофиза, надпочечников и половых желез. Это приводит к ретенционной азотемии и гиперазотурии. Нарушение промежуточного обмена приводит к снижению сопротивляемости инфекциям, тяжелым ангиопатиям.
В клинической стадии на передний план выступают полидипсия, полифагия, сухость слизистых ротовой полости, полиурия, ацетонурия, ацетонолактия, ацидоз, общая слабость, снижение и утрата продуктивных показателей, зудливость, сухость кожи, остеопороз, костно-суставная патология, изменения ЭКГ, протеинурия, ретинопатия, возможны гангрена конечностей, хвоста, нарушение пищеварения, признаки нарушения функций ЦНС, гипергликемическая кома.
Для диагностики редких форм сахарного диабета применяют исследование «сахарной кривой» - динамики уровня сахара в крови после сахарной нагрузки. Чем медленнее возвращается уровень гликемии к исходному показателю (до сахарной нагрузки), тем сильнее выражен сахарный диабет.
На картинке рядом с текстом представлено обобщенное описание эндокринных клеток островка Лангерганса , без указания их реальной позиции внутри него. На рисунке показана также структура фенестрированных капилляров и присутствующих в околокапиллярном пространстве автономных нервных волокон (HB) и нервных окончаний (НО).
| Статьи по теме: | |
|
Школьная энциклопедия К какой расе относятся сирийцы
90 % населения Сирии составляют мусульмане, 10% христиане. Мусульмане... Лимонный кекс на кефире с маком
Лимонный кекс на кефире без яиц (с пропиткой) — мой любимый рецепт к... Салат из сайры - простые и оригинальные рецепты аппетитной закуски Салат из сайры консервированной с рисом
Я очень люблю салаты с консервированной рыбой. Их можно готовить... | |