Выбор читателей




Популярные статьи
На планете Земля представлено огромное разнообразие национальностей, которым характерно определенное вероисповедание, традиции, культурные ценности. Более обширным понятием выступают расы, объединяющие людей по морфологическим признакам. Они сформировались в результате эволюции и социально-исторического развития населения. Расовая принадлежность человека всегда представляла интерес, ее происхождение, формирование, признаки изучает антропология.
Этимология слова «раса» появилась с середины XIX века в результате заимствования с французского языка «race», немецкого языка «rasse». Дальнейшая судьба слова неизвестна. Однако есть версия, что понятие произошло от латинского слова «generatio», что означает «способность к рождению».
Расой называют такую систему человеческих популяций, которая характеризуется сходством по наследственным биологическим признакам (внешнему фенотипу), что сформировались в определенной географической местности.
К морфологическим признакам, позволяющим делить население на группы, относят:
Не следует путать понятия национальности, нации и расы. Последнее может включать в себя представителей разных народностей и культур.
Значение рас заключается в формировании у населения приспособительных особенностей, которые облегчают существование на определенной территории. Изучением групп людей с идентичными морфологическими признаками занимается раздел антропологии - расоведение. Наука рассматривает определение, классификацию, как появились, факторы развития и формирования расовых особенностей.
До XX века число рас, сколько существует в мире, составляло 4, в зависимости от характерных признаков. Обширные группы объединяли представителей человечества, при этом различия во внешности зачастую становились причиной, почему между народами происходили распри и конфликты.
Основные расы людей, какие есть на земле с учетом территории расселения приведены в таблице:

За пределами Африканского континента негроидов нет. Австралоиды расположены в пределах определенного ареала. Процентное соотношение рас на земле распределилось по следующим показателям:
В Антарктиде жителей нет.
После XX века стала распространенной следующая классификация, которая включает 3 расовых типа. Это явление обусловлено объединением негроидной и австралоидной группы в смешанные расы.
Выделяют современные разновидности рас:
Расовое разделение включает в себя 2 ствола: западный и восточный.
К восточному стволу относятся американоиды, австралоиды и монголоиды. К американоидной расе по антропологическим признакам относятся индейцы.
Общепринятой классификации разделения по различным признакам нет, что считается прямым свидетельством непрерывности биологических процессов изменчивости.
Расовые особенности включают в себя множество характеристик строения человека, которые формируются под воздействием наследственного фактора и влияния окружающей среды. Внешние признаки человеческого облика изучает биология.
Расы интересовали специалистов с древних времен. Их отличительные особенности, описание, картинки, помогают понять расовую принадлежность конкретного человека.
Представители белых людей характеризуются светлым или смуглым оттенком кожи. Волосы прямые или волнистые от светлого до темного цвета. У мужчин на лице растет волосяной покров. Форма носа узкая выступающая, губы тонкой формы. К этой расе относятся .


Выделяют подрасы европеоидной расы:
Для первого вида характерны темные, а для второго - светлые волосы, глаза и кожа.
Облик классического европейца олицетворяет фальская раса. Фалиды выступают разновидностью расы кроманид, которая подверглась нордическому влиянию. Второе название этого подтипа - кроманид северный. От нордидов отличаются низким и широким лицом, низкой посаженной переносицей, выраженным красным оттенком кожи, крутой формой лба, короткой шеей и массивным туловищем.

Фалиды распространены в Нидерландах, Дании, Норвегии, Польше, Швеции, Исландии, Германии, западной части Прибалтики. В России фалиды встречаются редко.
Австралоиды включают в себя веддоидов, полинезийцев, айнцев, австралийцев и меланизийцев.

Выделяют несколько особенности австралоидной расы:

Сложно распознать представителя австралоидной расы ввиду смешения различных наций.
Люди монголоидной принадлежности имеют особые черты, позволяющие приспосабливаться к сложным климатическим условиям: пескам и ветрам в пустыне, снежным заносам.

К характеристикам монголоидной внешности относят ряд признаков:

Отличающим признаком выступает небольшой рост.
Желтокожие монголоиды преобладают по численности среди населения.

Для четвертой группы характерен список особенностей:

На лице волосяной покров не развит, борода и усы слабо выражены.
Продолжительный промежуток времени представителями высшей расы считались люди с белым цветом кожи. На почве этого развязывались военные конфликты в борьбе за первую расу на земле. За право господствовать на планете беспощадно истреблялись целые народы.
Отмечают некоторые интересные факты о происхождении рас. Немецкий антрополог Ф. Блуменбах считал наиболее красивыми представителями грузин. Присутствует специальный термин «кавказская раса», которая считается самой многочисленной.
Распространено смешение крови представителей разных групп. Например, мулат - термин, как называется смесь азиата и европейца. Смесь негроидной и монголоидной расы определяется самбо, а европеоида и монголоида - метис.

Интерес представляет вопрос, к какой расе относятся индусы - они образовались от австралоидной группы.

Расены выступают одной из известной разновидности Великой Расы. В мировой истории ее потомки назывались Тирренами.

Внешность расенов характеризуется рядом признаков:
Чаще всего у расенов 2 группа крови. Представителям этой расы характерны непоколебимость, сильный дух и ярость, что способствовало высокому уровню военной готовности.
Выступают восточнославянским этносом. По количеству это самый многочисленный народ на планете. По данным Википедии всего насчитывается 133 миллиона представителей русской национальности.

Расшифровка расизма: «Дискриминация людей по этническому происхождению, цвету кожи, культуре, гражданству, вероисповеданию и родному языку».
Термином называют реакционную идеологию и политику, которая направлена на оправданную эксплуатацию людей.
Расцвет расизма приходится на середину XIX века в Америке и Англии, Германии и Франции. Именно это послужило идеологической поддержкой торговли рабами, захвата земель колониями в Океании, Австралии, Азии, Африке, Америке.
Расисты придерживаются идеологии, что между психическими, интеллектуальными, социальными качествами и физическим строением есть определенная связь. Выделялись высшие и низшие расы.
Приверженцы расисткой идеологии считали, что первоначально возникали чистые расы, а позже смешение народов формировало новые. Дети появлялись с комбинированными особенностями внешности.
Считается, что метис отличается от своих кровных родителей:
Проблемой кровосмешения является кризис самоидентификации: во время военных конфликтов затруднено определение личности к одному гражданству и национальности.
Метисация отмечается постоянно и в результате на границах ареалов возникают переходные типы, сглаживающие различия.
Смешение рас с точки зрения науки рассматривается как видовое единство людей, их родство и плодовитость потомства. Однако проблемой выступает возможное исчезновение малочисленного народа или мелкой ветви большой расы.
Расизм противоречит идеалам любого общества людей. Он выступает глобальной проблемой человечества.
Европеоидная Монголоидная Негро-австралоидная
(евразийская) (азиатско-американская) (экваториальная)
Большие расовые стволы
Западный ствол Восточный ствол (европеоиды, негроиды, австралоиды) (монголоиды, американоиды)
4.1.Характеристика «больших» рас
а) Европеоиды
|
Морфологические признаки | |
|
1.Цвет кожи |
|
|
2. Окраска и форма волос и глаз |
|
|
3 . Форма губ и носа |
|
|
4. Волосяной покров |
|
|
5.Форма лица и подбородка |
|
|
7. Основное проживание |
*ортогнатизм (от греч. orthós - прямой и gnáthos - челюсть) - отсутствие или незначительность выступания вперёд верхней челюсти по отношению к общей фронтальной плоскости лица.
Европеоидная раса - раса, распространённая до эпохи Великих географических открытий в Европе, западной Азии и северной Африке, Индостан; позже - на всех населённых континентах, особенно широко европеоиды расселились в Северной Америке, Австралии, Северной Азии и Дальнем Востоке.
Европеоидная раса подразделена на три главные группы:
· Южная – со смуглой кожей, тёмными глазами и волосами
· Северная – со светлой кожей, значительной долей серых и голубых глаз, русых и белокурых волос
· Промежуточная – для которой характерна среднеинтенсивная пигментация
Характерные признаки
Распространённая ошибка - считать, что это «белые люди». На самом деле около миллиарда автохтонных европеоидов имеют чёрные глаза, волосы и смуглую кожу (например, население Индостана, Северной Африки). Отличается от других рас в первую очередь сильной профилировкой лица, густым волосяным покровом. Остальные признаки широко варьируют.
Расы северной группы: Атланто-балтийская, Беломорско-балтийская
Расы промежуточной (переходной) группы: Альпийская, Среднеевропейская, Восточноевропейская
Расы Южной группы: Средиземноморская, Индо-афганская, Балкано-кавказская, Переднеазиатская (Арменоидная), Памиро-ферганская

рис. 21 Представители Европеоидной расы
б) Монголоиды
|
Морфологические признаки | |
|
1.Цвет кожи |
|
|
2. Окраска и форма волос и глаз |
|
|
3 . Форма губ и носа |
|
|
4. Волосяной покров |
|
|
5.Форма лица и подбородка |
|
|
7. Основное проживание |
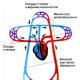
На Земле уже насчитывается около 6 млрд. людей. Среди них нет, и не
может быть двух совершенно одинаковых людей; даже близнецы, развившиеся из
одного яйца, несмотря на огромное сходство их внешнего облика, и
внутреннего строения, всегда какими-то мелкими чертами отличаются друг от
друга. Наука, изучающая изменения физического типа человека, известна под
именем «антропологии» (греч, «антропос» – человек). Особенно сильно заметны
телесные различия между территориальными группами людей, удаленными друг
от друга и живущими в различной естественно-географической обстановке.
Деление вида Homo Sapiens на расы произошло два с половиной столетия назад.
Происхождение термина «раса» точно не установлено; возможно, что он
представляет собой видоизменение арабского слова «рас» (голова, начало,
корень). Есть также мнение, что термин этот связан с итальянским razza, что
значит «племя». Слово «раса» примерно в том смысле, как оно употребляется
теперь, встречается уже у французского ученого Франсуа Бернье, который
Расы - это исторически сложившиеся группировки (группы популяции) людей
разной численности характеризующиеся сходством морфологических и физиологических свойств, а также общностью занимаемых ими территорий.
Развиваясь под влиянием исторических факторов и относясь к одному виду
(H.sapiens), раса отличается от народа, или этноса, который, обладая
определенной территорией расселения, может содержать несколько расовых
комплексов. К одной и той же расе может принадлежать ряд народов и
носителей многих языков. Большинство ученых сходятся во мнении, что
существует 3 крупных расы, которые в свою очередь распадаются на более
мелкие. В настоящее время по мнению разных ученых, насчитывается 34 – 40
рас. Расы отличаются друг от друга по 30-40 элементам. Расовые особенности
наследственны и являются приспособительными к условиям существования.
Целью моей работы является систематизировать и углубить знания о
человеческих расах.
Расы и их происхождение.
Наука о расах называется Расоведение. Расоведение изучает расовые
особенности (морфологические), происхождение, формирование, историю.
1.1. История человеческих рас.
О существовании рас люди знали еще до нашей эры. Тогда же были предприняты
и первые попытки, объяснить их происхождение. Например, в мифах древних
греков возникновение людей с черной кожей объяснялось неосторожностью сына
бога Гелиоса Фаэтона, который на солнечной колеснице так приблизился к
Земле, что обжег стоявших на ней белых людей. Греческие философы в
объяснениях причин возникновения рас большое значение придавали климату. В
соответствии с библейской историей родоначальниками белой, желтой и черной
рас были сыновья Ноя - любимый богом Яфет, Сим и проклятый богом Хам
соответственно.
Стремления систематизировать представления о физических типах народов,
населяющих земной шар, датируются XVII веком, когда, опираясь на различия
людей в строении лица, цвета кожи, волос, глаз, а также особенности языка и
культурных традиций, французский врач Ф. Бёрнье впервые в 1684 году
разделил человечество на {три расы - европеоидную, негроидную и
монголоидную). Сходную классификацию предлагал К.Линней, который, признавая
человечество в качестве единого вида, выделял дополнительную (четвертую)
pacy - лапландскую (население северных районов Швеции и Финляндии). В 1775
году Ж. Блюменбах разделил род человеческий на пять рас- кавказскую
(белую), монгольскую (желтую), эфиопскую (черную), американскую, (красную)
и малайскую (коричневую), а в 1889 году русский ученый И.Е.Деникер - на
шесть основных и более чем на двадцать дополнительных рас.
На основании результатов изучения антигенов крови (серологических
различий) У.Бойдом в 1953 году было выделено в человечестве пять рас.
Несмотря на наличие современных научных классификаций, в наше время очень
широко распространено подразделение человечества на европеоидов, негроидов,
монголоидов и австралоидов.
1.2. Гипотезы о происхождении рас.
Представления о происхождении рас и первичных очагах расообразования
отражены в нескольких гипотезах.
В соответствии с гипотезой полицентризма, или полифилии, автором которой
является Ф.Вайденрайх (1947), существовало четыре очага расообразования - в
Европе или Передней Азии, в Африке южнее Сахары, в Восточной Азии, в Юго-
Восточной Азии и на Больших Зондских островах. В Европе или Передней Азии
сложился очаг расообразования, где на основе европейских и переднеазиатских
неандертальцев возникли европеоиды. В Африке из африканских неандертальцев
образовались негроиды, в Восточной Азии синантропы дали начало монголоидам,
а в Юго-Восточной Азии и на Больших Зондских островах развитие
питекантропов и яванских неандертальцев привело к формированию
австралоидов. Следовательно, европеоиды, негроиды, монголоиды и австралоиды
имеют свои собственные очаги расообразования. Главным в расогенезе были
мутации и естественный отбор. Однако эта гипотеза вызывает возражения. Во-
первых, в эволюции не известны случаи, когда бы идентичные эволюционные
результаты воспроизводились несколько раз. Более того, эволюционные
изменения всегда новые. Во-вторых, научных данных о том, что каждая раса
обладает своим собственным очагом расообразования, не существует. В рамках
гипотезы полицентризма позднее Г.Ф.Дебец (1950) и Н.Тома (I960) предложили
два варианта происхождения рас. По первому варианту, очаг расообразования
европеоидов и африканских негроидов существовал в Передней Азии, тогда как
очаг расообразования монголоидов и австралоидов был приурочен к Восточной и
Юго-Восточной Азии. Европеоиды передвигались в пределах Европейского
материка и прилегающих к нему районов Передней Азии.
По второму варианту, европеоиды, африканские негроиды и австраловды
составляют один ствол расообразования, тогда как азиатские монголоиды и
американоиды- другой.
B соответствии с гипотезой моноцентризма, или. монофилии (Я.Я.Рогинский,
1949), которая основана на признании-общности происхождения, социально-
психического развития, а так же одинакового уровня физического и
умственного развития всех рас, последние возникли от одного предка, на
одной территории. Но последняя измерялась многими тысячами квадратных
километров Предполагается, что формирование рас произошло на территориях,
Восточного Средиземноморья, Передней и, возможно Южной Азии.
2. Механизм образования рас.
Выделяют четыре этапа расообразования (В.П.Алексеев, 1985) На первом
этапе имело место формирование первичных очагов расообразования
(территории, на которых происходит этот процесс) и основных расовых
стволов,-западного (европеоиды, негроиды и австралоиды) и восточного
(азиатские монголоиды и монголоиды и американоиды). Хронологически это
приходится на эпохи низшего или среднего палеолита (около 200 000 лет
назад), т.е. совпадает с возникновением человека современного типа.
Следовательно, основные расовые сочетания в западных и восточных районах
Старого Света складывались одновременно с оформлением признаков, присущих
современному человеку, а также с переселением части человечества в Новый
Свет. На втором этапе происходило выделение вторичных очагов
расообразования и формирование ветвей в пределах основных расовых стволов.
Хронологически этот этап приходится на верхний палеолит и частично мезолит
(около 15 000 - 20 000 лет назад).
На третьем этапе расообразования происходило становление локальных рас. По
времени это канун мезолита и неолита (около 10 000 - 12 000 лет назад).
На четвертом этапе возникли четвертичные очаги расообразования и
сформировались популяции с углубленной расовой дифференциацией, сходной
ссовременной. Это началось в эпоху бронзы и раннего железа, т.е. в IV-III
тысячелетиях до нашей эры.
2.1. Факторы расогенеза.
Среди факторов расогенеза наибольшая роль принадлежит естественному отбору,
особенно на ранних стадиях расообразования. За цвет кожи ответственны
кожные клетки, содержащие пигмент называемый меланином. Все люди, за
исключением альбиносов, имеют в клетках кожи меланин, количество которого
детерминируется генетически. В частности, образование пигмента определяется
присутствием гена, контролирующего тирозиназу которая катализирует
конверсию тирозина в меланин. Однако помимо тирозиназы на пигментацию кожи
оказывает влияние еще один фермент, за который ответствен другой ген,
меланина. Когда этот фермент синтезируется, то меланин образуется в малых
количествах и кожа является белой. Напротив, когда он отсутствует (не
синтезируется), то меланин образуется в больших количествах и кожа является
значение и меланин, стимулирующий гормон. Таким образом, в контроле цвета
кожи принимают участие, по меньшей мере, три пары генов.
Важность цвета кожи в качестве расового признака объясняется связью между
солнечным освещением и продукцией витамина D, который необходим для
поддержания в организме баланса кальция. Избыток этого витамина
сопровождается отложением кальция в костях и ведет к их хрупкости, тогда
как недостаток кальция имеет следствием рахит. Между тем количество
витамина D, синтезируемого в норме, контролируется дозой солнечного
облучения, которое проникает в клетки, находящиеся глубже слоя меланина.
Чем больше меланина в коже, тем меньше света он пропускает. До периода,
когда были разработаны способы искусственного обогащения пищи витамином D,
люди были зависимы в продукции витамина D от солнечного света. Чтобы
витамин D синтезировался в оптимальных количествах, т.е. достаточных для
поддержания нормального баланса кальция, люди со светлой кожей должны были
обитать на определенной географической широте далеко от экватора, где
солнечная радиация более слабая. Напротив, люди с черной кожей должны были
находиться ближе к экватору. Как видно, территориальное распределение людей
с разной пигментацией кожи является функцией географической широты.
Просветление кожи у европеоидов способствует проникновению солнечного света
глубоко в ткани человека, что ускоряет синтез противорахитического витамина
D, который обычно медленно синтезируется в условиях недостатка солнечной
радиации. Миграции людей с интенсивно пигментированной кожей к отдаленным
от экватора широтам, а людей с недостаточно пигментированной кожей - к
тропическим широтам могла вести к дефициту витамина D у первых и избытку у
вторых с вытекающими последствиями. Таким образом, в прошлом цвет кожи имел
селективное значение для естественного отбора.
Значительно выступающий узкий нос у европеоидов удлиняет носоглоточный
путь, благодаря чему холодный воздух нагревается, что защищает от
переохлаждения гортань и легкие. Развитие слизистых способствует большей
теплоотдаче. Курчавые волосы лучше предохраняют голову от перегрева, так
как создают воздухоносную прослойку. Вытянутая высокая голова тоже
нагревается слабее, чем широкая и низкая. Несомненно, что эти признаки
являются адаптивными. Таким образом, в результате мутаций и естественного
отбора многие расовые признаки возникали как приспособление к условиям
географической среды обитания.
К факторам расогенеза относятся также дрейф генов, изоляция и смешение
популяций.
Дрейф генов, контролирующих признаки, способен изменять генетическую
структуру популяций. Подсчитано, что в результате дрейфа генов облик
популяций может измениться на протяжении 50 поколений, т.е. около 1250 лет.
Сущность генетического дрейфа состоит в том, что в изолированных
популяциях, где почти все браки эндогамны, значительно повышаются шансы
встречи в аллельных парах рецессивных генов, снижается уровень
гетерозиготности и повышается концентрация рецессивов в гомозиготном
состоянии.
В популяциях (демах), где на протяжении многих поколений браки заключаются
преимущественно внутри своей группы, с течением времени могут происходить
заметные изменения расовых особенностей, которые приводят к тому, что
первоначально сходные популяции оказываются различными. Возникновение
таких различий, не носящих адаптивного характера, является результатом
сдвигов в частоте появления отдельных признаков. Они приводят к тому, что
одни особенности могут совершенно исчезнуть, а другие получить очень
широкое распространение.
Изоляция популяций проявлялась в различных формах и объемах. Например,
географическая изоляция первобытных коллективов в эпоху палеолита
сопровождалась дифференциацией их генетического состава, прерыванием
контактов с другими коллективами. Различные географические барьеры
оказывали влияние не только на генетическую дифференциацию популяций, но и
на концентрацию культурных традиций.
Смешение популяций имело значение в далеком прошлом и еще большее значение
При формировании молодых рас. В далеком прошлом более прогрессивные формы
встречались с архаичными, что вело не только к истреблению последних, но и
к метисации. Среди «молодых» рас наиболее характерной является
североамериканская цветная раса (негритянское население США), которая
возникла в результате смешения лесной негроидной расы с расами банту, а
также северо-западной европейской, альпийской, средиземноморской и,
возможно, другими расами Южно-африканская цветная раса возникла на основе
банту, бушменов и готтентотов. В настоящее время на Гавайских островах
формируется новая расовая группа вследствие метисации европеоидов,
монголоидов и полинезийцев.
На современном этапе будущее рас определяется рядом действующих в наше
время факторов. Продолжается рост численности населения земного шара,
увеличивается его миграция, повышается частота межрасовых браков.
Предполагают, что благодаря двум последним факторам в отдаленном будущем
начнет формироваться единая раса человечества. В то же время возможно и
другое последствие межрасовых браков, связанное с формированием новых
популяций с собственными специфическими сочетаниями генов.
2.2 Роль условий среды на формирование рас.
Влияние природных условий на развитие человеческих рас, несомненно.
Первоначально в древнем человечестве оно было, наверное, более сильным, в
процессе же формирования современных рас сказывалось слабее, хотя и до сих
пор в некоторых при знаках, например в пигментации кожного покрова,
обнаруживается с достаточной ясностью. Влияние всей сложной совокупности
условий жизни, очевидно, имело важнейшее значение для возникновения,
формирования, ослабления и даже исчезновения расовых признаков.
При расселении по Земле люди попадали в разные природные условия. Но эти
условия, столь сильно влияющие на виды и подвиды животных, не могли так же
и с той же интенсивностью действовать на качественно отличные от них расы
человечества, все более и более использующего природу и преобразующего ее в
процессе общественного труда.
В эволюции разных человеческих групп многие расовые особенности,
несомненно, имели известное приспособительное значение, но позже в
значительной мере утратили его в связи с нарастанием роли факторов
общественного характера и постепенным ослаблением, и почти полным
прекращением действия естественного отбора. Первоначально большое значение
для развития рас имело расселение по новым областям, благодаря чему многие
группы людей, попав в разные природные условия, длительно существовали
отдельно друг от друга. Образ питания их соответственно дифференцировался.
Позже, однако, по мере увеличения численности человечества, все более
усиливалось соприкосновение расовых групп, которое приводило к процессу их
смешения друг с другом.
3. Расогенез и генетика.
Раньше существовало представление, согласно которому каждый индивидум
несет признаки определенной расы. Оно получило название типологической
концепции расы. Это название связано с вполне четкой и практически лишь
единственной задачей расового анализа - определением расового типа
индивидуума. В основе типологической концепции расы лежала гипотеза
наследования расовых признаков, согласно которой они передаются от
поколения к поколению целым комплексом. Это означает, что расовые свойства
наследственно сцеплены, гены расовых признаков расположены в одной или
нескольких близких хромосомах и любой расовый признак физиологически
неразрывно связан со всеми остальными. Но физиологическая зависимость между
расовыми признаками на самом деле либо совсем отсутствует, либо очень
слаба. На что указывают низкие коэффициенты корреляции между расовыми
признаками. Существует группа ученых, которая утверждает о независимом
наследовании расовых признаков, их первый основной постулат - индивидуум не
есть носитель расовых свойств. Второй постулат - популяция и раса (как
группа популяций) представляет собой не сумму, а совокупность индивидуумов;
внутри популяции и расы действуют определенные закономерности изменчивости.
Расовая изменчивость – групповая, а не индивидуальная, и о ней имеет смысл
говорить, начиная с популяционного уровня. Сходные морфологически и
генетически популяции, образующие ту или иную расовую общность, связаны
между собой не случайно, а в силу происхождения или каких-то иных
исторических причин. Раса, любая расовая общность состоит из отдельных
исторически организованных элементов, но это, однако, не индивидуумы (как
думали раньше), а популяции. Мозаика расовой изменчивости складывается из
мозаики популяционной изменчивости. Обе они в совокупности и создают все
богатство изменчивости человеческого вида. Каждую популяцию стали изучать
не как сумму индивидуумов, а как неповторимое по своей специфике сочетание
групповых свойств. Популяционная концепция опирается на достижения
популяционной генетики, новейшей биометрии, математической теории эволюции
Большое количество материала накоплено в антропологии по теме происхождения рас на Земле. Основные расы - это европеоидная, монголоидная, австралоидная, негроидная. Остальные либо менее распространенные, либо смешанного происхождения. Не будем вдаваться в подробности морфологических отличий, это не цель данного исследования, а только попробуем установить прямую связь между данными антропологии и генетики в этом вопросе.Европеоидная раса характеризуется светлой кожей (с вариациями от очень светлой, главным образом в Северной Европе, до относительно смуглой в Южной Европе), мягкими прямыми или волнистыми волосами, горизонтальным разрезом глаз, умеренно или сильно развитым волосяным покровом на лице и груди у мужчин, заметно выступающим носом, прямым или несколько наклонным лбом.
Известно, что основная масса большой европеоидной расы
имела исходный ареал где-то в пределах обширной области, охватывавшей некоторые районы
юго-западной Азии, а также южной Европы и северной Африки - это всё районы Средиземноморья
. Само происхождение европеоидов вероятно связано с еще
менее обширным ареалом на Ближнем Востоке около 40 тыс. лет назад. Т.к. формирование европеоидной расы связано с целым каскадом мутаций, вероятность
повторения которого в нескольких группах независимо на практике исключена - это и целый комплекс мутаций ответственных за пигментацию кожи, волос, глаз; и за строение носа,
формы черепа, челюстей и т.д.
На основе расчета возрастов для основных гаплогрупп в ДНК-генеалогии, а также предполагаемых мест возникновения, можно указать единственную гаплогруппу, достоверно
относящуюся к большой европеоидной расе на момент возникновения - это гаплогруппа IJ (снип M429). Её возраст на основе расчета скоростей мутаций составляет примерно те самые 40 тыс. лет, а место возникновения - Ближний Восток. От тех носителей IJ, что мигрировали в северном направлении через несколько тысячелетий образовалась гаплогруппа I, а от тех что мигрировали в районе западной Азии - гаплогруппа J. Вероятно, на самых ранних этапах формирования европеоидной расы
были и другие гаплогруппы в очаге формирования, но они не оставили потомков по прямой мужской линии. На самых ранних этапах в зоне формирования европеоидной расы присутствовала и гаплогруппа G (снип M201),
отделившаяся от F вероятно в восточной части территории Ближнего Востока.
Гаплогруппа F, как родительская для IJK, G, H имеет возраст не менее 55 тыс. лет и потому группа
носителей с этим маркером (снип M89) скорее всего обладала либо каким-то недифференцированным набором морфологических признаков, либо была еще разнородна и не формировала более-менее единого антропологического типа. Какие-то отдельные представители IJK могли остаться на территории Ближнего Востока наравне с IJ и G и далее мигрировать на
северо-запад и юго-запад, но большая часть гаплогруппы IJK направилась на восток, где примерно через 8 тыс. лет произошла мутация, определяющая гаплогруппы K, H. Произошло это в районе Южной
Азии. Через несколько тысяч лет при миграции далее на восток от K возникли новые гаплогруппы - это LT (снип P326), которая на первых порах так и осталась в Южной Азии, а от тех
представителей K, что мигрировали в юго-восточную Азию возникла гаплогруппа MNOPS, она же KxLT, определяемая снипом M526. Гаплогруппа MNOPS возкла около 40 тыс. лет назад в районе современного п-ва
Индокитай, находящегося в юго-восточной Азии. От данной гаплогруппы в дальнейшем отделилась сначала гаплогруппа P (снип M45), ушедшая на запад, в южную Азию. Потом
гаплогруппы M и S, что мигрировали на юг и юго-восток, в Полинезию и в Папуа-Новую Гвинею. И наконец гаплогруппа NO (снип M214), что мигрировала на восток и север.
Гаплогруппа P при миграции на север образовала новую гаплогруппу Q (снип M242), что была одной из первых, заселивших север Азии и в дальнейшем переправившихся около 13 тыс. лет назад на Американский континент. Другой потомок P - это гаплогруппа R (снип M207), возникшая в Южной Азии в дальнейшем мигрировала на север (в Сибирь), где образовались субклады R1 (снип M173) и R1a (снип M420).
Среди мт-ДНК гаплогрупп, характерных для того же региона Ближнего Востока и участвовавших в формировании европеоидов на начальном этапе вероятно относились гаплогруппы UK, HV и JT, I. Все они значительно распространены в Европе, северной Африке и на Ближнем Востоке. Это и были спутницы европейских кроманьонцев - носителей Y-гаплогруппы IJ. Предположительный возраст для гаплогруппы U составляет более 55 тыс. лет, что можно объяснить значительно меньшим отбором для мт-ДНК линий, вследствие более лояльного отношения к женщинам. Для мужских линий характерно прохождение через "бутылочные горлышки", что омолаживает возраст общего предка и разнообразие Y-линий.
В аутосомном сравнении популяций еще более полная картина распространения европеоидов
и прото-европеоидов, по которой наибольшее количество экспансий пришлось на неолит. Это были
миграции в основном с Ближнего Востока первых земледельческих и скотоводческих культур. (Современное распространение аутосомных компонент представлено в исследовании
генетики европейцев).
Многими антропологами предполагается, что миграции
прото-европеоидов в палеолите около 20-30 тыс. лет назад на восток могли доходить отдельными группами вплоть до Южной Азии на юге и Алтая на севере - можно
констатировать, что эти группы были либо малочисленны относительно основного неевропеоидного населения и просто "растворились" в них, либо они мирным
или не мирным путем были ассимилированы более восточными группами прото-монголоидов. После получения результатов аутосомной дДНК со стоянки Мальта в Сибири (неподалеку от озера Байкал) стало ясно, что аутосомная компонента, характерная для европеоидов у палеолитических жителей Сибири представлена была, но её процент и соотношение с другими "неевропеоидными" аутосомными компонентами южной Азии и Сибири, позволяют достаточно уверенно сказать, что прото-европеоидами они не были, об этом же говорят низкое переносье и сравнительно малый изгиб чешуи лобной кости без надбровного валика у фрагмента с Афонтовой горы, хотя аутосомный профиль останков с Афонтовой горы (недалеко от Красноярска) несколько ближе к современным Восточно-Европейцам. ДНК останков со стоянки Мальта по прямым линиям относятся к мт-ДНК гаплогруппе U, а по Y-хромосоме к гаплогруппе R.
Сегодня на севере Азии, вплоть до Алтая на юге, распространена мт-ДНК гаплогруппа U4, которая также может быть остаточным следом древних прото-европеоидов Сибири и вероятно какой-то из линий гаплогруппы I*; для Южной Азии это предположительно мт-ДНК гаплогруппа U2. Прямых Y-линий от тех мигрантов не осталось, но предположительно это могли быть либо какие-то субклады гаплогруппы G, либо IJ. Учитывая что возраст мт-ДНК гаплогруппы U составляет не менее 55 тыс. лет, то некоторые субклады данной гаплогруппы могли относиться к недифференцированному расовому типу, близкому прото-европеоидам или прото-монголоидам - возможно U2 и U4 были их характерными "мито-метками".
Карта верхнепалеолитических миграций Y-гаплогрупп и субкладов
Алексей Зоррин
Характеризуется темным цветом кожи и глаз, волновыми или кудрявыми волосами, широким носом, который мало выступает, поперечным расположением ноздрей, полными губами и определенными отличными признаками черепа. Борода и усы вырастают едва заметно. Все перечисленные признаки носят адаптивный характер. Например, темная кожа меньше повреждается лучами солнца, препятствует проникновению ультрафиолетовых лучей в кожу, оберегает ее от ожогов. Кудрявые волосы создают плотную войлочную шапку, которая надежно защищает голову от излучений солнца.
Характеризуется смугловатой или светлой кожей, прямым, чаще твердыми волосами, средней шириной губ и носа, сплющенной лица, сильно выступающими скулами, сравнительно большим размером лица, заметным развитием «третьего века». Наиболее важными адаптивными признаками монголоидной расы является отложение жировой клетчатки на лице у детей, узкие щели глаз, складка века. Все эти признаки в прошлом имели приспособительное значение от обморожения, ветра, пыли и солнечного света, который отражался от снега.
Характеризуется светлой или смугловатой кожей, прямыми или волновыми мягкими волосами, хорошо развитым волосяным покровом на лице у мужчин (борода и усы), узким выступающим носом, тонкими губами, а также определенными отличными признаками черепа. Наиболее важным приспособительным признаком этой расы является большой размер носовой полости, что связано с необходимостью создания своеобразной «камеры, которая нагревает» холодный воздух.
| Статьи по теме: | |
|
Журнал осмотра детей на чесотку и педикулез — форма и оформление Журнал осмотра контактных детей
Отрасль: Образование Специализации: Заведующему детским садом,... Это (,) наверное (,) вводные слова
Выражение «на самом деле» запятой может отделяться от остального... Несколько способов гадания на свадьбу: все карты Вам в руки
Для любой девушки очень волнительным и важным вопросом считается... | |