Выбор читателей




Популярные статьи
В полости рта липиды подвергаются лишь механической обработке. В желудке имеется небольшое количество липазы, которая гидролизует жиры. Малая активность липазы желудочного сока связана с кислой реакцией содержимого желудка. Кроме того, липаза может влиять только на эмульгированные жиры, в желудке отсутствуют условия для образования эмульсии жира. Только у детей и у моногастричных животных липаза желудочного сока играет важную роль в переваривании липидов.
Кишечник является основным местом переваривания липидов. В двенадцатиперстной кишке на липиды воздействует желчь печени и сок поджелудочной железы, одновременно происходит нейтрализация кишечного содержимого (химуса). Происходит эмульгирование жиров под действием желчных кислот. В состав желчи входят: холевая кислота, дезоксихолевая (3,12 дигидроксихолановая), хенодезоксихолевая (3,7 дигидроксихолановая) кислоты, натриевые соли парных желчных кислот: гликохолевая, гликодезоксихолевая, таурохолевая, тауродезоксихолевая. Они состоят из двух компонентов: холевой и дезоксихолевой кислот, а также глицина и таурина.
дезоксихолевая кислота хенодезоксихолевая кислота
гликохолевая кислота
таурохолевая кислота
Соли желчных кислот хорошо эмульгируют жиры. При этом увеличивается площадь соприкосновения ферментов с жирами и увеличивается действие фермента. Недостаточность синтеза желчных кислот или задержка поступления нарушает эффективность действия ферментов. Жиры, как правило, всасываются после гидролиза, но часть тонко эмульгированных жиров всасывается через стенку кишечника и переходит в лимфу без гидролиза.
Эстеразы разрывают в жирах эфирную связь между, спиртовой группой и карбоксильной группой карбоновых кислот и неорганических кислот (липаза, фосфатазы).
Под действием липазы жиры гидролизуются на глицерин и высшие жирные кислоты. Активность липазы возрастает под действием желчи, т.е. желчь непосредственно активирует липазу. Кроме того, активность липазы увеличивают ионы Са ++ вследствие того, что ионы Са ++ образуют нерастворимые соли (мыла) с освободившимися жирными кислотами и предотвращают их подавляющее влияние на активность липазы.
Под действием липазы в начале гидролизуются эфирные связи у α и α 1 (боковых) углеродных атомов глицерина, затем у β-углеродного атома:
Под действием липазы до 40% триацилглицеридов расщепляются до глицерина и жирных кислот, 50-55% гидролизуется до 2-моноацилглицеринов и 3-10% не гидролизуется и всасываются в виде триацилглицеринов.
Стериды корма расщепляются ферментом холестеролэстеразой до холестерина и высших жирных кислот. Фосфатиды гидролизуются под влиянием фосфолипаз А, A 2 , С и D. Каждый фермент действует на определенную сложноэфирную связь липида. Точки приложения фосфолипаз представлены на схеме:
Фосфолипазы поджелудочной железы, тканевые фосфолипазы вырабатываются в виде проферментов и активируются трипсином. Фосфолипаза A 2 змеиных ядов катализирует отщепление ненасыщенной жирной кислоты в положении 2 фосфоглицеридов. При этом образуются лизолецитины с гемолитическим действием.
фосфотидилхолин лизолецитин
Поэтому при попадании этого яда в кровь происходит сильный гемолиз.. В кишечнике эта опасность устраняется действием фосфолипазы A 1 , быстро инактивирующей лизофосфатид в результате отщепления от него остатка насыщенной жирной кислоты с превращением его в неактивный глицерофосфохолин.
Лизолецитины в малых концентрациях стимулируют дифференцировку лимфоидных клеток, активность протеинкиназы С, усиливают клеточную пролиферацию.
Коламинфосфатиды и серинфосфатиды расщепляются фосфолипазой А до лизоколаминфосфатидов, лизосеринфосфатидов, которые далее расщепляются фосфолипазой A 2 . Фосфолипазы С и D гидролизуют связи холина; коламина и серина с фосфорной кислотой и остатка фосфорной кислоты с глицерином.
Всасывание липидов происходит в тонком отделе кишечника. Жирные кислоты с длиной цепи менее 10 углеродных атомов всасываются в неэтерифицированной форме. Для всасывания необходимо присутствие эмульгирующих веществ – желчных кислот и желчи.
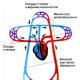
Ресинтез жира, характерного для данного организма, происходит в кишечной стенке. Концентрация липидов в крови в течение 3-5 часов после приема корма высокая. Хиломикроны – мелкие частицы жира, образующиеся после всасывания в кишечной стенке, представляют собой липопротеиды, окруженные фосфолипидами и белковой оболочкой, внутри содержат молекулы жира и желчных кислот. Они поступают в печень, где липиды подвергаются промежуточному обмену, а желчные кислоты проходят в желчный пузырь и далее обратно в кишечник (см. рис.9.3 на стр.192). В результате такого кругооборота теряется малое количество желчных кислот. Считают, что молекула желчной кислоты в сутки совершает 4 кругооборота.
Несомненно, что в повседневной пище из жиров доминируют нейтральные жиры, известные как триглицериды, каждая молекула которых включает глицериновое ядро и боковые цепи, состоящие из трех жирных кислот. Нейтральные жиры - основной компонент животной пищи, а в растительной пище их содержится крайне мало.
В обычной пище имеется небольшое количество фосфолипидов, холестерола и эфиров холестерола. Фосфолипиды и эфиры холестерола содержат жирные кислоты и, следовательно, могут рассматриваться как жиры. Впрочем, холестерол является представителем стеринов и не содержит жирных кислот, но проявляет некоторые физические и химические свойства жиров; к тому же он производится из жиров и легко превращается в них. Следовательно, с диетологической точки зрения, холестерол рассматривается как жир.
Переваривание жиров в кишечнике . Небольшое количество триглицеридов переваривается в желудке под действием лингвальной липазы, которая секретируется железами языка в ротовой полости и проглатывается вместе со слюной. Количество перевариваемых таким образом жиров составляет менее 10%, а потому не существенно. Основное переваривание жиров происходит в тонком кишечнике, о чем сказано далее.
Эмульгирование жиров желчными кислотами и лецитином. Первый этап переваривания жиров заключается в физическом разрушении капель жира на мелкие частицы, поскольку водорастворимые ферменты могут действовать только на поверхности капли. Этот процесс называют эмульгированием жиров, он начинается в желудке с перемешивания жиров с другими продуктами переваривания желудочного содержимого.
Далее основной этап эмульгирования происходит в двенадцатиперстной кишке под влиянием желчи, секрета печени, который не содержит пищеварительных ферментов. Однако желчь содержит большое количество желчных солей, а также фосфолипид - лецитин. Эти компоненты, в особенности лецитин, чрезвычайно важны для эмульгирования жиров. Полярные частицы (места, в которых происходит ионизация воды) желчных солей и молекул лецитина хорошо растворимы в воде, тогда как большая оставшаяся часть этих молекул хорошо растворима в жирах.
Таким образом, жирорастворимые порции секрета печени растворяются в поверхностном слое жировых капель вместе с выступающей полярной частью. В свою очередь, выступающая полярная часть растворима в окружающей водной фазе, что значительно снижает поверхностное натяжение жиров и делает их также растворимыми.
Когда поверхностное натяжение капли нерастворимой жидкости низкое, нерастворимая в воде жидкость во время перемещения значительно легче разрушается на множество мелких частиц, чем при более высоком поверхностном натяжении. Следовательно, основная функция желчных солей и лецитина - делать капли жира способными к легкому размельчению при перемешивании с водой в тонком кишечнике. Это действие аналогично действию синтетических моющих средств, широко используемых в домашнем хозяйстве для устранения жира.
Каждый раз в результате перемешивания в тонком кишечнике диаметр жировых капель существенно уменьшается, поэтому общая жировая поверхность многократно увеличивается. Из-за того, что средний диаметр частичек жира в кишечнике после эмульгирования оказывается менее 1 мкм, общая площадь жировой поверхности, образованная в результате процесса эмульгирования, увеличивается в 1000 раз.
Фермент липаза является водорастворимым и может воздействовать только на поверхности жировых капель. Отсюда понятно, насколько значительна детергентная роль лецитина и желчных солей в переваривании жиров.
Спасибо
Сайт предоставляет справочную информацию исключительно для ознакомления. Диагностику и лечение заболеваний нужно проходить под наблюдением специалиста. У всех препаратов имеются противопоказания. Консультация специалиста обязательна!
 Липиды
представляют собой одну из групп органических соединений, имеющую огромное значение для живых организмов. По химической структуре все липиды делятся на простые и сложные. Молекула простых липидов состоит из спирта и желчных кислот, в то время как в состав сложных липидов входят и другие атомы или соединения.
Липиды
представляют собой одну из групп органических соединений, имеющую огромное значение для живых организмов. По химической структуре все липиды делятся на простые и сложные. Молекула простых липидов состоит из спирта и желчных кислот, в то время как в состав сложных липидов входят и другие атомы или соединения.В целом, липиды имеют огромное значение для человека. Эти вещества входят в значительную часть продуктов питания , используются в медицине и фармации, играют важную роль во многих отраслях промышленности. В живом организме липиды в том или ином виде входят в состав всех клеток. С точки зрения питания – это очень важный источник энергии.
 Липиды входят в состав практически всех тканей организма. Их молекулы есть в любой живой клетке, и без этих веществ попросту невозможна жизнь. В организме человека встречается очень много различных липидов. Каждый вид или класс этих соединений имеет свои функции. От нормального поступления и образования липидов зависит множество биологических процессов.
Липиды входят в состав практически всех тканей организма. Их молекулы есть в любой живой клетке, и без этих веществ попросту невозможна жизнь. В организме человека встречается очень много различных липидов. Каждый вид или класс этих соединений имеет свои функции. От нормального поступления и образования липидов зависит множество биологических процессов.С точки зрения биохимии, липиды принимают участие в следующих важнейших процессах:
В живом организме липиды выполняют следующие функции:
Также жировая ткань, отложенная в подкожно-жировой клетчатке, осуществляет теплоизоляцию. Ткани, богатые липидами, в целом хуже проводят тепло. Это позволяет организму поддерживать постоянную температуру тела и не так быстро охлаждаться или перегреваться в различных условиях внешней среды.
Таким образом, сами липиды не выполняют регуляторной функции, но их недостаток может отразиться на многих процессах в организме.
Также липиды в той или иной степени связаны с обменом следующих веществ:
Липиды нерастворимы в воде, поэтому в двенадцатиперстной кишке они не сразу подвергаются расщеплению ферментами. Сначала происходит так называемое эмульгирование жиров. После этого химические связи расщепляются под действием липазы, поступающей из поджелудочной железы. В принципе, для каждого вида липидов сейчас определен свой фермент, отвечающий за расщепление и усвоение данного вещества. Например, фосфолипаза расщепляет фосфолипиды, холестеролэстераза – соединения холестерола и т. д. Все эти ферменты в том или ином количестве содержатся в соке поджелудочной железы.
Расщепленные фрагменты липидов всасываются по отдельности клетками тонкого кишечника. В целом переваривание жиров представляет собой весьма сложный процесс, который регулируется множеством гормонов и гормоноподобных веществ.
Процесс эмульгирования липидов в пищеварительной системе проходит в несколько этапов:
За расщепление липидов отвечают следующие группы ферментов:
Наибольшую роль в усвоении и поддержании постоянного уровня липидов играют следующие вещества:
Биосинтез липидов происходит в следующих тканях и клетках:
На первом этапе ресинтез липидов происходит в стенках кишечника. Здесь поступающие с пищей жирные кислоты преобразуются в транспортные формы, которые отправятся с кровью в печень и другие органы. Часть ресинтезированных липидов будет доставлено в ткани, из другой части образуются необходимые для жизнедеятельности вещества (липопротеины, желчь, гормоны и др. ), избыток преобразуется в жировую ткань и откладывается «про запас».
В состав миелиновой оболочки в нервной системе входят следующие липиды:
Липиды входят в состав следующих жизненно важных гормонов:
Для волос и кожи липиды важны по следующим причинам:
 В биологии и химии существует довольно много различных классификаций липидов. Основной является химическая классификация, согласно которой липиды делятся в зависимости от своей структуры. С этой точки зрения все липиды можно разделить на простые (состоящие только из атомов кислорода, водорода и углерода
) и сложные (включающие хотя бы один атом других элементов
). Каждая из этих групп имеет соответствующие подгруппы. Эта классификация наиболее удобна, так как отражает не только химическое строение веществ, но и частично определяет химические свойства.
В биологии и химии существует довольно много различных классификаций липидов. Основной является химическая классификация, согласно которой липиды делятся в зависимости от своей структуры. С этой точки зрения все липиды можно разделить на простые (состоящие только из атомов кислорода, водорода и углерода
) и сложные (включающие хотя бы один атом других элементов
). Каждая из этих групп имеет соответствующие подгруппы. Эта классификация наиболее удобна, так как отражает не только химическое строение веществ, но и частично определяет химические свойства.В биологии и медицине имеются свои дополнительные классификации, использующие другие критерии.
После попадания в организм все экзогенные липиды расщепляются и усваиваются живыми клетками. Здесь из их структурных компонентов будут сформированы другие липидные соединения, в которых нуждается организм. Эти липиды, синтезированные собственными клетками, называются эндогенными. Они могут иметь совершенно другую структуру и функции, но состоят из тех же «структурных компонентов», которые попали в организм с экзогенными липидами. Именно поэтому при недостатке в пище тех или иных видов жиров могут развиваться различные заболевания. Часть компонентов сложных липидов не может быть синтезирована организмом самостоятельно, что отражается на течении определенных биологических процессов.
В природе жирные кислоты содержатся в самых разных веществах - от нефти до растительных масел. В организм человека они попадают в основном с пищей. Каждая кислота является структурным компонентом для определенных клеток, ферментов или соединений. После всасывания организм преобразует ее и использует в различных биологических процессах.
Наиболее важными источниками жирных кислот для человека являются:
Ненасыщенные жирные кислоты делятся на две большие группы:
Жировая ткань в организме человека представлена в основном триглицеридами. Большая часть этих веществ, перед тем как отложиться в жировой ткани, претерпевает некоторые химические трансформации в печени.
①. ЖЕЛУДОК : у взрослого человека в желудке пищевые триглицериды практически не расщепляются, т.к.
рН ↓ под действием НСl до 1,5 (опт. рН для липазы = 5,5-7,5). Также в желудке отсутствуют
условия для эмульгирования , а липаза может действовать только на триглицериды , находящиеся в
форме эмульсии );
у грудных детей в желудке (рН〜5,2) под действием лингвальной липазы происходит активно
гидролиз эмульгированных жиров молока.
②. КИШЕЧНИК : рН под действием гидрокарбонатов , растворенных в щелочном содержимом сока
поджелудочной железы и желчи .
В составе сока поджелудочной железы в кишечник поступают:
1). липаза – гидролиз жиров до свободных жирных кислот и моноацилглицеролов , может
отщеплять жирную кислоту по 1 -положению глицерина в фосфолипидах ;
1). холестеролэстераза поджелудочной железы – гидролиз пищевых эфиров холестерина;
2). фосфолипаза А 2 – отщепляет жирную кислоту от фосфолипидов по 2 -положению глицерина ;
3). щелочная фосфатаза кишечного сока – гидролиз по фосфоэфирным связям фосфолипидов .
Всасывание холестерола , частично расщепленных фосфолипидов происходит с помощью мицелл,
формируемых солями парных желчных кислот .
ОРНИТИНОВЫЙ ЦИКЛ МОЧЕВИНООБРАЗОВАНИЯ (ОЦ ) (в печени) - основной путь детоксикации аммиака (N Н 3 ), который образуется при распаде азот -содержащих веществ: аминокислот, биогенных аминов, пуринов и пиримидинов, фосфо- и гликолипидов, гексозаминов, гликозаминогликанов, гема и др. Реакции ОЦ направлены на связывание токсичного Аммиака с образованием нетоксичной Мочевины . Азот в мочевине происходит из карбамоил-фосфата (N Н 3 +СО 2 ) и аспарагиновой кислоты .
В ОЦ участвуют а/к – орнитин (непротеиногенная), аспартат и образуется аргинин ).
(На образование 1 молекулы Мочевины расходуется энергия 3 АТФ , которые ресинтезируются за счет превращения промежуточного метаболита ОЦ – фумарата через малат в ОА (в ц. Кребса), что сопровождается восстановлением НАДНН + , обеспечивающим синтез 2,5 АТФ в ПДЦМХ . Образованная молекула ОА в реакции трансаминирования при участии пиридоксаминфосфата (кофермент В 6 ) превращается в аспартат , аминогруппа которого, наряду с молекулой аммиака , используется в синтезе Мочевины (N Н 2 -СО -N Н 2 )).
Гипераммониемия (аммиака в крови) – при ↓ активности ферментов ОЦ . Проявления: учащенное дыхание, возбудимость, мигрень, судороги, рвота при употреблении белковой пищи.
ТРАНСДЕЗАМИНИРОВАНИЕ (НЕПРЯМОЕ ДЕЗАМИНИРОВАНИЕ – НД ) - процесс дезаминирования α-аминокислот (а/к ) с образованием α-кетокислот (к/к ) без промежуточного освобождения аммиака . Протекает НД в 2 этапа : 1 - трансаминирование , катализируемое В 6 -зависимой аминотрансферазой : происходит перенос N Н 2 –группы с а/к на α-кетоглутарат , с образованием к/к и глутамата , соответственно. Витамин В 6 вступает в реакцию в форме кофермента – пиридоксаль-Ф , который принимает от а/к амино группу и превращается в пиридоксамин-Ф (через образование промежуточных шиффовых оснований – альдимин и кетимин ), который далее отдает N Н 2 –группу на α-кетоглутарат с образованием глутамата . 2 – окислительное дезаминирование глутамата при участии глутаматдегидрогеназы с выделением аммиака и образованием α-кетоглутарата . Другие типы дезаминирования : восстановительное, гидролитическое (у м/орг.), внутримолекулярное (гистидин → урокановая к-та).
ТРАНСРЕАМИНИРОВАНИЕ (НЕПРЯМОЕ АМИНИРОВАНИЕ – НА ) - процесс, обратный непрямому дезаминированию , обеспечивающий связывание аммиака с образованием из α-к/к → α-а/к . НА протекает в 2 этапа: 1 – восстановительное аминирование α–кетоглутарата с образованием глутамата и 2 – трансаминирование : перенос аминогруппы с глутамата на α-к/к с образованием α-а/к .
БИОГЕННЫЕ АМИНЫ (БА ) – биологически активные производные аминокислот , ключевой реакцией образования которых является В 6 –зависимое декарбоксилирование а/к . К БА относятся: ГАМК – декарбоксилированное производное глутамата ; Гистамин – декарбоксилированный гистидин , Серотонин – образуется из триптофана (при В 6 –зав. декарбоксилировании и гидроксилировании при участии вит.С ); Катехоламины : Дофамин , Норадреналин , Адреналин – образуются из тирозина (при участии В 6 -зависимой декарбоксилазы, вит.С -зависимой гидроксилазы, S АМ -зависимой метилтрансферазы).
ГЛИКО- и КЕТОГЕННЫЕ АМИНОКИСЛОТЫ : Гликогенные а/к – а/к, которые, распадаясь, превращаются в ЩУК и ПВК, а далее через 3 -й обходной путь вступают в глюконеогенез → далее в гликогеногенез . Кетогенные а/к (ЛЛИФТТ ) – а/к, при распаде которых образуется ацетоацетат (кетоновое тело ) или ацетилКоА (при концентрации которого синтезируются кетоновые тела ). Лиз, Лей – строго кетогенные а/к ; И/лей, Ф/а, Тир, Трп – смешанные – глико - и кетогенные а/к.
НЕЗАМЕНИМЫЕ АМИНОКИСЛОТЫ : Вал, Лей, Изолей, Мет, Ф/а, Трп, Тре, Лиз. ; Полузаменимые – Тир, Цист .; Частично заменимая – Арг. ; Незаменимая в детском возрасте – Гист .
ЗНАЧЕНИЕ АМНОКИСЛОТ : ① . ФЕНИЛАЛАНИН (незам.а/к) → ТИРОЗИН (полузамен. а/к) – глико- и кетогенные а/к являются предшественниками: 1 ). катехоламинов (в мозговом веществе надпочечников, в мозге) – дофамин , норадреналин , адреналин ; 2 ). йодтиронинов (в щитовидной железе) – трийодтиронин (Т 3 ), тетрайодтиронин (Т 4 ); 3 ). пигмента меланина (в коже, волосах, радужке).
Фенилкетонурия (выведение фенилпирувата с мочой) – наследственная энзимопатия , связанная с ↓ активности фермента – фенилаланинмонооксигеназы , гидроксилирующей фенилаланин в тирозин . Проявления: олигофрения , возбудимость, мышечная гипотония.
Алкаптонурия (выведение алкаптона с мочой) – наследственная энзимопатия , связанная с ↓ активности фермента – гомогентизатдиокигеназы , участвующего в обмене тирозина , что сопровождается гомогентизиновой кислоты и образованием из нее черного пигмента – алкаптона . Проявления: алкаптон откладывается в суставах , что сопровождается воспалением и ограничением их подвижности; развивается охроноз , связанный с отложением алкаптона в хрящах ушных раковин и крыльев носа.
Альбинизм – развивается при нарушении синтеза меланина из тирозина . Отмечается депигментация радужки глаз, волос; чувствительность кожи к УФ.
Гипотиреоз –↓ выработки Т 3 и Т 4 , что приводит к кретинизму в детском возрасте и микседеме (слизистый отек) – у взрослых. Гипертиреоз - выработки Т 3 и Т 4 , что приводит к развитию Базедовой болезни (развиваются экзофтальм, зоб, тахикардия, t 0). (Ферменты , обеспечивающие синтез Т 3 и Т 4 , из тирозина : 1) - йодидпероксидаза , активирующая пищевой йод ; 2) - тирозинйодиназа , включающая йод по С 3 и С 5 -положениям тирозина с конденсацией 2 -х молекул тирозина ).
②. ТРИПТОФАН (незам.; глюко- и кетогенная а/к) необходим для синтеза: 1 ). серотонина (биогенный амин) – регулирует многие соматические функции организма и является антидепрессантом; 2 ). мелатонина – гормон эпифиза, регулирующий биоритмы; 3 ). витамина РР (НАД + , НАДФ + ) в печени.
Гиповитаминоз В 6 – ↓ -сяактивность В 6 – зависимой кинурениназы и нарушается обмен триптофана , что сопровождается выведением с мочой побочного метаболита – ксантуреновой кислоты , и нарушением синтеза витамина РР (НАД + , НАДФ + ). Развивается пеллагроподобный дерматит.
Болезнь «голубых пеленок» - связана с нарушением обмена триптофана , что сопровождается образования индолилацетата , индикана (окрашивают пеленки новорожденных в голубой цвет). Проявления: пеллагроподобный дерматит, эмоциональная лабильность, атаксия, запоры.
③. МЕТИОНИН (незам. а/к ) и ЦИСТЕИН (полузамен. а/к ) – серосодержащие а/к: 1 ). Метионин в форме кофермента SAM участвует в синтезе: а). адреналина из норадреналина ; б). мелатонина из серотонина , в). холина из этаноламина (холин входит в состав лецитина , ацетилхолина ), г). креатин-фосфата (наряду с арг и гли ) – мышечный макроэрг, д). карнитина (наряду с лиз ) – переносчик ЖК через мембраны МХ, е). полиаминов – спермина , спермидина (наряду с орнитином ) – регулируют процессы клеточного роста и дифференцировки, активируя синтез ДНК, РНК, белка, ж). ансерина из карнозина (наряду с гист и β-ала ) – повышают амплитуду мышечного сокращения в утомленной мышце.
2 ). ЦИСТЕИН – необходим для синтеза: а). тиоэтиламина , который участвует в образовании из витамина Пантотеновая кислота коферментов – КоА S Н и 4-фосфопантотеина , б). глутатиона – трипептида, включающего также глу и гли – участвует в переносе а/к через мембраны, в восстановлении дегидроаскорбиновой кислоты , в инактивации активных форм кислорода , в восстановлении SH - групп ферментов и мембран э/ц, в). таурина – образует парные желчные кислоты (таурохолевая, таурохенодезоксихолевая ).
Гомоцистеинурия - выведение с мочой гомоцистеина , т.к. ↓ активность цистатионин-синтазы . Нарушается синтез цис , ↓ умственное развитие, судороги, остеопороз (↓ гидроксилирование лиз в коллагене ), дрожание радужки глаз.
Цистинурия -экскреции с мочой цистеина , цистина при нарушении почечной реабсорбции. Образуются цистиновые камни, происходит закупорка мочевыводящих путей.
④. ЛИЗИН (н/з ), АРГИНИН (частично н/з а/к ), ГИСТИДИН (н/з в детском возрасте ) – оснóвные а/к, «+ » - заряж. ЛИЗИН и АРГИНИН : 1 ).входят в состав гистонов (Н1, Н2а, Н2в, Н3, Н4 ), 2 ). ЛИЗИН – участвует в преобразовании вит.Н в кофермент – биоцитин ; Липоевой кислоты – в липамид ; участвует в организации активного центра аминотрансфераз , связывая коферменты В 6 . 3 ). АРГИНИН – используется для синтеза креатин-фосфата (наряду с гли и мет ), орнитина . Образуется арг в орнитиновом цикле . 4 ). ГИСТИДИН – является предшественником гистамина ; участвует в образовании карнозина и ансерина ; в связывании гемоглобином О 2 .
⑤. ГЛУТАМАТ, АСПАРТАТ (замен. а/к ) – дикарбоновые а/к , «-»-заряж. – участвуют: 1 ). в связывании аммиака с образованием глутамина и аспарагина , 2 ). в синтезе пуринов и пиримидинов , 3 ). в образовании альбуминов и глобулинов крови, 4 ). в трансаминировании , 5 ). АСПАРТАТ – участвует в орнитиновом цикле , 6). ГЛУТАМАТ – в синтезе ГАМК , глутатиона .
⑥. ГЛИЦИН (зам. а/к ) - участвует в синтезе: 1 ). глутатиона (наряду с цис и глу ), 2 ). гема (наряду с СукцинилКоА ), 3 ). пуринов – аденина , гуанина , 4 ). парных желчных кислот – гликохолевая , гликохенодезоксихолевая , 5 ). креатин-фосфата (наряду с арг и мет ), 6 ). участвует в детоксикации продуктов гниения белков , 7 ). в образовании активной формы ТГФК (кофермент фолиевой кислоты ) – N 5 , N 10 -метилен-ТГФК , необходимой для синтеза серина , тимина из урацила .
ПРОСТЫЕ (ПБ ) и СЛОЖНЫЕ БЕЛКИ (СБ ): ПБ – протеины : состоят только из аминокислот (альбумины, глобулины, гистоны, протамины, проламины ). СБ – протеиды : состоят из апопротеина и простетической небелковой группы (металлопротеины (трансферрин, церулоплазмин ), фосфо- , нуклео- (РНК-содерж. - рибосома , ДНК-содерж. - нуклеосома ), хромо- (цветные белки : Нв, Мв, цитохромы , родопсин, флавопротеиды ), глико- , липопротеины ).
Фибриллярные белки – коллаген , эластин ; фибронектин и ламинин (адгезивные белки ); кератины волос; актин и миозин – сократительные белки; фиброин шелка и паутины.
Азотистый баланс – соотношение количества азота , поступающего в организм в составе пищи и выделяемого с мочой, потом, калом. При положительном АБ – происходит задержка азота в организме – в растущем организме, при беременности, при восстановлении после болезни. Отрицательный АБ – больше азота выводится (гиперазотурия ) – при активном распаде тканевых белков при гипертиреозе, сахарном диабете, распаде злокачественной опухоли.
ПРОДУКТЫ ГНИЕНИЯ БЕЛКОВ (ПГБ ) и ПУТИ ИХ ДЕТОКСИКАЦИИ : ПГБ – скатол ,индол (образуются из трп ), крезол ,фенол (из ф/а и тир ),сероводород (из цист , мет ),кадаверин (из лиз )и др. - образуются в результате разложения пищевых белков и а/к микрофлорой нижних отделов кишечника. Усиливаются процессы гниения при ↓ протеолитической функции поджелудочной железы (остр., хронич. панкреатит) Основное место детоксикации ПГБ – печень , где происходит их метилирование, ацилирование, конъюгирование с глюкуроновой и серной кислотами , с глицином .
По гиппуровой кислоте в моче (продукт конденсации бензойной кислоты с глицином ) судят о детоксикационной функции печени .
ОБМЕН НУКЛЕОТИДОВ (Н/Т ): Н/Т - состоят из азотистого основания (АО ) (пуриновые АО – аденин и гуанин , пиримидиновые – тимин , урацил , цитозин) , рибозы/дезоксирибозы и остатка фосфорной кислоты (ФК ).
1 . при распаде Н/Т : отщепляется ФК под действием нуклеотидаз → далее отщепляется пентоза → происходит дезаминирование АО (кроме урацила ) и превращение АО в конечные продукты : для пуринов – мочевая кислота (МК ) (предшественники – гипоксантин , ксантин ), для пиримидинов – β-аланин (для тимина – β-аминоизомасляная кислота ). Нуклеозиды отличаются от н/т – отсутствием ФК .
Подагра – отложение в суставах плохо растворимых Nа-солей мочевой кислоты при гиперурикемии (концентрации МК в крови), что является результатом употребления пурин -содержащих продуктов (кофе, икра), или ↓ активности гуанин , гипоксантин-фосфорибозил-трансферазы . Развиваются боли в суставах, позвоночнике, ↓ их подвижность, гиперурикемия , уратурия ( МК и ее Nа-солей в моче).
Болезнь Леша-Найхана – наследственная энзимопатия , связанная со ↓ гуанин,гипоксантин-фосфорибозилтрансферазы . Проявления: возбудимости, ↓ умственного развития, нанесение самоповреждений, почечно-каменная болезнь.
Гиперурикемия, уратурия – мочевой кислоты в крови и моче при подагре, болезни Леша-Найхана, патологии почек, печени, при лейкемии.
2 . синтез Н/Т : а ). ПУРИНОВЫЕ Н/Т синтезируются за счет формирования пуринового кольца на активированной фосфорибозе – фосфорибозилпирофосфате (Ф R РР ) из глицина , аспартата , 2 -х молекул глутамина , и при участии формил- и метен- ТГФК (кофермент фолиевой кислоты – В 9 ). При этом образуется Инозиновая кислота , которая далее аминируется с образованием АМФ (источник аминогруппы – аспартат ) и ГМФ (источник аминогруппы – глутамин ). б ). При синтезе ПИРИМИДИНОВЫХ Н/Т сначала формируется Оротовая кислота (из карбамоил-фосфата и аспартата ), которая далее переносится на Ф R РР с образованием оротидилмонофосфата (ОМФ ). ОМФ , декарбоксилируясь , превращается в УМФ , который при участии глутамина → в ЦМФ , а при участии метилен-ТГФК – в ТМФ .
Значение Н/Т : являются мономерами ДНК и РНК ; АТФ – универсальный макроэрг ; выполняют коферментную функцию. Значение УТФ и ЦТФ как коферментов : УТФ – участвует в синтезе гликогена , во взаимопревращении галактозы в глюкозу , в синтезе гликолипидов , гликозаминогликанов . ЦТФ – участвует в синтезе фосфолипидов .
Оротацидурия – выведение оротовой кислоты с мочой при ↓ активности ОМФ-декарбоксилазы , что сопровождается ↓ синтеза пиримидинов – урацила , тимина , цитозина (нарушается пролиферация и дифференцировка быстро делящихся клеток). Развивается мегалобластическая анемия, дерматиты.
СИНТЕЗ ГЕМА : из Глицина и СукцинилКоА образуется Аминолевулиновая кислота , 2 -е молекулы которой формируют порфобилиноген , из 4 -х молекул которого синтезируется уропорфириноген (УПГ ), который через n- количество стадий превращается в Протопорфирин- IX , в который феррохелатаза встраивает железо (Fе 2+ ) и образуется ГЕМ . Значение ГЕМА - является простетической группой хромопротеидов (цветные белки): Нв , Мв, цитохромы ; является коферментом каталазы и пероксидазы .
Порфирии (эритропоэтическая, печеночная, кожная и др.) – наследственные энзимопатии , связанные со ↓ активности какого-либо фермента, участвующего в синтезе гема (напр., ↓активности уропорфириноген- III -синтазы ). Развиваются гипертрихоз, фотодерматит, эритродонтия; с мочой выводятся порфобилиноген , уропорфириноген и др. промежуточные метаболиты.
РАСПАД ГЕМА : под действием гем-окисляющей системы ГЕМ последовательно превращается в вердоглобин → биливердин → билирубин , который в крови адсорбируется на альбуминах , превращаясь в непрямой билирубин (НБ , 75 %) . В печени , конъюгируясь с глюкуроновой или серной кислотами (при участии глюкуронил- и ФАФС – фосфоаденозинфосфат–сульфо трансферазы ), происходит образование прямого билирубина (ПБ – нетоксичный, растворимый, дает прямую реакцию с диазореактивом ) . Поступая в кишечник билирубин многократно восстанавливается и превращается в стеркобилин – конечный продукт распада гема , который в норме выводится с калом (300 мг ) и мочой (2-3 мг ).
Желтухи – развиваются при уровня билирубина в крови (гипербилирубинемия ). Различают 3 типа желтух : 1 – гемолитическая (при гемолиза эритроцитов): Н Б в крови, стеркобилин в кале и моче; 2 – паренхиматозная (при гепатитах, циррозах): появление уробилиногена в моче (предшественник стеркобилина ), в крови уровня общего БР ; 3 – обтурационная , механическая (при закупорке желчных протоков камнем, опухолью): ПБ в крови, отсутствует стеркобилин в кале и моче (ахоличный – бесцветный кал), ПБ выводится с мочой (билирубинурия ) – моча приобретает цвет «темного пива».
СИНТЕЗ БЕЛКА : 1. транскрипция – переписывание последовательности нуклеотидов (н/т ) ДНК в последовательность н/т и РНК по принципу комплементарности (между пуринами и пиримидинами : А=Т (У ), Г ≡Ц ), с заменой Т на У . 2. посттранкрипционный процессинг – созревание про- и РНК : вырезание интронов , сплайсинг – сшивание экзонов , «кэпирование» и РНК по 5′-концу: (+)-е метилированных н/т ), присоединение полиаденилата по 3′-концу. 3. трансляция (происходит на рибосомах при участии т РНК , приносящей а/к к месту синтеза полипептидной цепи) – раскодирование последовательности н/т и РНК в последовательность аминокислот белка : 3 н/т кодируют 1 а/к (триплетность генетического кода ). Свойства генетического кода : универсальность, триплетность, вырожденность, неперекрываемость. 4. посттрансляцинный процессинг (фолдинг – процесс сворачивания белка в правильную пространственную биологически активную конформацию при участии белков-шаперонов ; присоединение простетической группы в сложных белках).
Процессы транскрипции , трансляции , репликации протекают в 3 этапа : инициация, элонгация, терминация.
РЕПЛИКАЦИЯ ДНК : удвоение ДНК (при делении клетки) происходит при участии следующих ферментов: 1. хеликаза - раскручивает двойную спираль ДНК с образованием репликативной вилки , 2. топоизомераза – предупреждает суперспирализацию ДНК в местах формирования репликативной вилки , 3. праймаза – катализирует образование «затравочного» праймера (олиго рибо нуклеотид ), с которого начинается синтез ДНК , 4. ДНК-полимераза III (основной фермент репликации , катализирующий синтез лидирующей цепи ДНК и отстающей цепи – фрагментами Оказаки в направлении 5′→ 3′), ДНК-полимераза I (удаляет затравочный праймер и замещает на олиго дезоксирибо нуклеотид ),ДНК-полимераза II (участвует в репарации – устранении ошибок); 5. ДНК-лигаза (сшивает фрагменты Оказаки , соединяет 2 цепи ДНК ).

Роль липидов в питании
Липиды являются обязательной составной частью сбалансированного пищевого рациона человека. Принято считать, что при сбалансированном питании соотношение белков, липидов и углеводов в пищевом рационе составляет примерно 1: 1: 4. В среднем в организм взрослого человека с пищей ежесуточно поступает около 80 г жиров животного и растительного происхождения. В пожилом возрасте, а также при малой физической нагрузке потребность в жирах снижается, в условиях холодного климата и при тяжелой физической работе - увеличивается.
Значение жиров как пищевого продукта весьма многообразно. Прежде всего жиры в питании человека имеют важное энергетическое значение. Высокая калорийность жиров по сравнению с белками и углеводами придает им особую пищевую ценность при расходовании организмом больших количеств энергии. Известно, что 1 г жиров при окислении в организме дает 38,9 кДж (9,3 ккал), тогда как 1 г белка или углеводов - 17,2 кДж (4,1 ккал). Следует также помнить, что жиры являются растворителями витаминов A, D, Е и др., в связи с чем обеспеченность организма этими витаминами в значительной степени зависит от поступления жиров в составе пищи. Кроме того, с жирами в организм вводятся некоторые полиненасыщенные кислоты (линолевая, линоленовая, арахидоновая), которые относят к категории незаменимых жирных кислот, ибо ткани человека и ряда животных потеряли способность синтезировать их. Эти кислоты условно объединены в группу под названием "витамин F".
Наконец, с жирами организм получает комплекс биологически активных веществ, таких, как фосфолипиды, стерины и др., играющих важную роль в обмене веществ.
Переваривание и всасывание липидов
Расщепление жиров в желудочно-кишечном тракте. Слюна не содержит расщепляющих жиры ферментов. Следовательно, в полости рта жиры не подвергаются никаким изменениям. У взрослых людей жиры проходят через желудок также без особых изменений, поскольку содержащаяся в небольшом количестве в желудочном соке взрослого человека и млекопитающих липаза малоактивна. Величина pH желудочного сока около 1,5, а оптимальное значение pH для желудочной липазы находится в пределах 5,5-7,5. Кроме того, липаза может активно гидролизовать только предварительно эмульгированные жиры, в желудке же отсутствуют условия для эмульгирования жиров.
Переваривание жиров в полости желудка играет важную роль в процессе пищеварения у детей, особенно грудного возраста. Известно, что pH желудочного сока у детей грудного возраста около 5,0, что способствует перевариванию эмульгированного жира молока желудочной липазой. К тому же есть основания полагать, что при длительном употреблении молока в качестве основного продукта питания у детей грудного возраста наблюдается адаптивное усиление синтеза желудочной липазы.
Хотя в желудке взрослого человека не происходит заметного переваривания жиров пищи, все же в желудке отмечается частичное разрушение липопротеидных комплексов мембран клеток пищи, что делает жиры более доступными для последующего воздействия на них липазы панкреатического сока. Кроме того, незначительное расщепление жиров в желудке приводит к появлению свободных жирных кислот, которые, поступая в кишечник, способствуют эмульгированию там жиров.
Расщепление жиров, входящих в состав пищи, происходит у человека и млекопитающих преимущественно в верхних отделах тонкого кишечника, где имеются весьма благоприятные условия для эмульгирования жиров.

После того как химус попадает в двенадцатиперстную кишку, здесь прежде всего происходит нейтрализация соляной кислоты желудочного сока, попавшей в кишечник с пищей, бикарбонатами, содержащимися в панкреатическом и кишечном соках. Выделяющиеся при разложении бикарбонатов пузырьки углекислого газа способствуют хорошему перемешиванию пищевой кашицы с пищеварительными соками. Одновременно начинается эмульгирование жира. Наиболее мощное эмульгирующее действие на жиры, несомненно, оказывают соли желчных кислот, попадающие в двенадцатиперстную кишку с желчью в виде натриевых солей, большая часть которых конъюгирована с глицином или таурином. Желчные кислоты представляют собой основной конечный продукт обмена холестерина.
Главные стадии образования из холестерина желчных кислот, в частности холевой кислоты, можно представить в следующем виде. Процесс начинается с гидроксилирования холестерина в 7-м α-положении, т. е. с включения гидроксильной группы в положении 7 и образования 7-гидроксихолестерина. Затем через ряд стадий образуется 3,7,12-тригидроксикопростановая кислота, боковая цепь которой подвергается β-окислению. В завершающей стадии отделяется пропионовая кислота (в виде пропионил-КоА) и боковая цепь укорачивается. Во всех этих реакциях принимает участие большое количество ферментов и коферментов печени.
По своей химической природе желчные кислоты являются производными холановой кислоты. В желчи человека в основном содержится холевая (3,7,12-триоксихолановая), дезоксихолевая (3,12-дигидроксихолано- и хенодеэоксихолевая (3,7-дигидроксихолановая) кислоты.
Кроме того, в желчи человека в малых (следовых) количествах содержится литохолевая (3-гидроксихолановая) кислота, а также аллохолевая и уреодезоксихолевая кислоты - стереоизомеры холевой и хенодезоксихолевой кислот.
Как уже отмечалось, желчные кислоты присутствуют в желчи в конъюгированной форме, т. е. в виде гликохолевой, гликодезоксихолевой, гликохенодезоксихолевой (около 2/3-4/3 всех желчных кислот) или таурохолевой, тауродезоксихолевой и таурохенодезоксихолевой (около 1/5-1/3 всех желчных кислот). Эти соединения иногда называют парными, так как они состоят из двух компонентов - из желчной кислоты и глицина или же желчной кислоты и таурина.
Заметим, что соотношения между конъюгатами этих двух видов могут меняться в зависимости от характера пищи: в случае преобладания в ней углеводов увеличивается относительнее содержание глициновых конъюгатов, а при высокобелковой диете - тауриновых конъюгатов. Строение этих конъюгатов может быть представлено в следующем виде:

Считается, что только комбинация: соль желчной кислоты + ненасыщенная жирная кислота + моноглицерид способна дать необходимую степень эмульгирования жира. Соли желчных кислот резко уменьшают поверхностное натяжение на поверхности раздела жир/вода, благодаря чему они не только облегчают эмульгирование, но и стабилизируют уже образовавшуюся эмульсию.
Желчные кислоты выполняют также важную роль в качестве своеобразного активатора панкреатической липазы 1 , под влиянием которой происходит расщепление жира в кишечнике. Вырабатываемая в поджелудочной железе липаза расщепляет триглицериды, находящиеся в эмульгированном состоянии. Считают, что активирующее влияние желчных кислот на липазу выражается в смещении оптимума действия данного фермента с pH 8,0 до 6,0, т. е. до той величины pH, которая более постоянно поддерживается в двенадцатиперстной кишке в ходе переваривания жирной пищи. Конкретный же механизм активации липазы желчными кислотами пока неясен.
1 Однако существует мнение, что активация липазы происходит не под влиянием желчных кислот. В соке поджелудочной железы присутствует предшественник липазы, который активируется в просвете кишки путем образования комплекса с колипазой (кофактором) в молярном соотношении 2: 1. Это способствует сдвигу оптимума pH с 9,0 до 6,0 и предотвращению денатурации фермента. Установлено также, что на скорость катализируемого липазой гидролиза не оказывает существенного влияния ни степень ненасыщенности жирных кислот, ни длина углеводородной цепи (от С 12 до С 18). Ионы кальция ускоряют гидролиз главным образом потому, что они образуют нерастворимые мыла с освобождающимися жирными кислотами, т. е. практически сдвигают реакцию в направлении гидролиза.

Есть основания считать, что существует панкреатическая липаза двух типов: одна из них специфична в отношении эфирных связей в положениях 1 и 3 триглицерида, а другая - гидролизует связи в положении 2. Полный гидролиз триглицеридов происходит постадийно: сначала быстро гидролизуются связи 1 и 3, а потом уже медленно идет гидролиз 2-моноглицерида (схема).
Необходимо отметить, что в расщеплении жиров участвует также кишечная липаза, однако активность ее невысока. К тому, же эта липаза катализирует гидролитическое расщепление моноглицеридов и не действует на ди- и триглицериды. Таким образом, практически основными продуктами, образующимися в кишечнике при расщеплении пищевых жиров, являются жирные кислоты, моноглицериды и глицерин.
Всасывание жиров в кишечнике . Всасывание происходит в проксимальной части тонкого кишечника. Тонко эмульгированные жиры (величина жировых капелек эмульсии не должна превышать 0,5 мкм) частично могут всасываться через стенку кишечника без предварительного гидролиза. Однако основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты, моноглицериды и глицерин. Жирные кислоты с короткой углеродной цепью (менее 10 С-атомов) и глицерин, будучи хорошо растворимыми в воде, свободно всасываются в кишечнике и поступают в кровь воротной вены, оттуда - в печень, минуя какие-либо превращения в кишечной стенке. Сложнее дело обстоит с жирными кислотами с длинной углеродной цепью и моноглицеридами. Всасывание этих соединений происходит при участии желчи и главным образом желчных кислот, входящих в ее состав. В желчи соли желчных кислот, фосфолипиды и холестерин содержатся в соотношении 12,5:2,5:1,0. Жирные кислоты с длинной цепью и моноглицериды в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы (мицеллярный раствор). Структура этих мицелл такова, что их гидрофобное ядро (жирные кислоты, глицериды и др.) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфолипидов. Мицеллы примерно в 100 раз меньше самых мелких эмульгированных жировых капель. В составе мицелл высшие жирные кислоты и моноглицериды переносятся с места гидролиза жиров к всасывающей поверхности кишечного эпителия. Относительно механизма всасывания жировых мицелл единого мнения нет. Одни исследователи считают, что в результате так называемой мицеллярной диффузии, а возможно и пиноцитоза, мицеллы целой частицей проникают в эпителиальные клетки ворсинок. Здесь происходит распад жировых мицелл; при этом желчные кислоты сразу же поступают в ток крови и через систему воротной вены попадают в печень, откуда они вновь секретируются в составе желчи. Другие исследователи допускают возможность перехода в клетки ворсинок только липидного компонента жировых мицелл. А соли желчных кислот, выполнив свою физиологическую роль, остаются в просвете кишечника. И лишь потом в подавляющем большинстве они всасываются в кровь (в подвздошной кишке), попадают в печень и затем выделяются с желчью. Таким образом, и те и другие исследователи признают, что происходит постоянная циркуляция желчных кислот между печенью и кишечником. Этот процесс получил название печеночно-кишечной (энтерогепатической) циркуляции.
С помощью метода меченых атомов было показано, что в желчи содержится лишь небольшая часть желчных кислот (10-15% от общего количества), вновь синтезированных печенью, т. е. основная масса желчных кислот желчи (85-90%)- это желчные кислоты, реабсорбированные в кишечнике и повторно секретируемые в составе желчи. Установлено, что у человека общий пул желчных кислот - примерно 2,8-3,5 г; при этом они совершают 5-6 оборотов в сутки.
Ресинтез жиров в стенке кишечника . В стенке кишечника синтезируются жиры, в значительной степени специфичные для данного вида животного и отличающиеся по своей природе от пищевого жира. В известной мере это обеспечивается тем, что в синтезе триглицеридов (а также фосфолипидов) в кишечной стенке принимают участие наряду с экзогенными и эндогенные жирные кислоты. Однако способность к осуществлению в станке кишечника синтеза жира, специфичного для данного вида животного, все же ограничена. А. Н. Лебедевым показано, что при скармливании животному, особенно предварительно голодавшему, больших количеств чужеродного жира (например, льняного масла или верблюжьего жира) часть его обнаруживается в жировых тканях животного в неизмененном виде. Жировые депо скорее всего являются единственной тканью, где могут откладываться чужеродные жиры. Липиды, входящие в состав протоплазмы клеток других органов и тканей, отличаются высокой специфичностью, их состав и свойства мало зависят от пищевых жиров.
Механизм ресинтеза триглицеридов в клетках стенки кишечника в общих чертах сводится к следующему: первоначально из жирных кислот образуется их активная форма - ацил-КоА, после чего происходит ацилирование моноглицеридов с образованием сначала диглицеридов, а затем триглицеридов:

Таким образом, в клетках кишечного эпителия высших животных моноглицериды, образующиеся в кишечнике при переваривании пищи, могут ацилироваться непосредственно, без промежуточных стадий.
Однако в эпителиальных клетках тонкого кишечника содержатся ферменты - моноглицеридлипаза, расщепляющая моноглицерид на глицерин и жирную кислоту, и глицеролкиназа, способная превращать глицерин (образовавшийся из моноглицерида или всосавшийся из кишечника) в глицерол-3-фосфат. Последний, взаимодействуя с активной формой жирной кислоты - ацил-КоА, дает фосфатидную кислоту, которая затем используется для ресинтеза триглицеридов и особенно глицерофосфолипидов (подробно см. ниже).
Переваривание и всасывание глицерофосфолипидов и холестерина . Вводимые с пищей глицерофосфолипиды подвергаются в кишечнике воздействию специфических гидролитических ферментов, разрывающих эфирные связи между компонентами, входящими в состав фосфолипидов. Принято считать, что в пищеварительном тракте распад глицерофосфолипидов происходит при участии фосфолипаз, выделяемых с панкреатическим соком. Ниже приведена схема гидролитического расщепления фосфатидилхолина:

Различают несколько типов фосфолипаз.
Фосфолипаза А 2 поджелудочной железы поступает в полость тонкого кишечника в неактивной форме и только после воздействия трипсина, приводящего к отщеплению от нее гептапептида, приобретает активность. Накопление лизофосфолипидов в кишечнике может быть устранено, если одновременно на глицерофосфолипиды действуют обе фосфолипазы: А 1 и А 2 . В результате образуется нетоксичный для организма продукт (например, при расщеплении фосфотидилхолина - глицерилфосфорилхолин).
Итак, в результате действия фосфолипаз глицерофосфолипиды расщепляются с образованием глицерина, высших жирных кислот, азотистого основания и фосфорной кислоты.
Необходимо отметить, что подобный механизм расщепления глицерофосфолипидов существует и в тканях организма; катализируется этот процесс тканевыми фосфолипазами. Заметим, что последовательность реакций расщепления глицерофосфолипидов на отдельные компоненты еще неизвестна.
Механизм всасывания высших жирных кислот и глицерина нами был уже рассмотрен. Фосфорная кислота всасывается кишечной стенкой главным образом в виде натриевых или калиевых солей. Азотистые основания (холин и этаноламин) всасываются в виде своих активных форм.
Как уже отмечалось, в кишечной стенке происходит ресинтез глицерофосфолипидов. Необходимые компоненты для синтеза: высшие жирные кислоты, глицерин, фосфорная кислота, органические азотистые основания (холин или этаноламин) поступают в эпителиальную клетку при всасывании из полости кишечника, поскольку они образуются при гидролизе пищевых жиров и липидов; частично эти компоненты доставляются в эпителиальные клетки кишечника с током крови из других тканей. Ресинтез глицерофосфолипидов идет через стадию образования фосфатидной кислоты.
Что касается холестерина, то он попадает в пищеварительные органы человека преимущественно с яичным желтком, мясом, печенью, мозгом. В организм взрослого человека ежедневно поступает 0,1-0,3 г холестерина, содержащегося в пищевых продуктах либо в виде свободного холестерина, либо в виде его эфиров (холестеридов). Эфиры холестерина расщепляются на холестерин и жирные кислоты при участии особого фермента панкреатического и кишечного соков - холестеролэстеразы. Нерастворимый в воде холестерин, подобно жирным кислотам, всасывается в кишечнике лишь в присутствии желчных кислот.
Образование хиломикронов и транспорт липидов . Ресинтезированные в эпителиальных клетках кишечника триглицериды и фосфолипиды, а также поступивший в эти клетки из полости кишечника холестерин (здесь он может частично этерифицироваться) соединяются с небольшим количеством белка и образуют относительно стабильные комплексные частицы - хиломикроны (ХМ). Последние содержат около 2% белка, 7% фосфолипидов, 8% холестерина и его эфиров и свыше 80% триглицеридов. Диаметр ХМ колеблется от 100 до 5000 нм. Благодаря большим размерам частиц ХМ не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и диффундируют в лимфатическую систему кишечника, а из нее - в грудной лимфатический проток. Затем из грудного лимфатического протока ХМ попадают в кровяное русло, т. е. с их помощью осуществляется транспорт экзогенных триглицеридов, холестерина и частично фосфолипидов из кишечника через лимфатическую систему в кровь. Уже через 1-2 ч после приема пищи, содержащей липиды, наблюдается алиментарная гиперлипемия. Это физиологическое явление, характеризующееся в первую очередь повышением концентрации триглицеридов в крови и появлением в ней ХМ. Пик алиментарной гиперлипемии приходится на 4-6 ч после приема жирной пищи. Обычно через 10-12 ч после приема пищи содержание триглицеридов возвращается к нормальным величинам, а ХМ полностью исчезают из кровяного русла.
Известно, что печень и жировая ткань играют наиболее существенную роль в дальнейшей судьбе ХМ. Последние свободно диффундируют из плазмы крови в межклеточные пространства печени (синусоиды). Допускается, что гидролиз триглицеридов ХМ происходит как внутри печеночных клеток, так и на их поверхности. Что же касается жировой ткани, то хиломикроны не способны (из-за своих размеров) проникать в ее клетки. В связи с этим триглицериды ХМ подвергаются гидролизу на поверхности эндотелия капилляров жировой ткани при участии фермента липопротеидлипазы, который тесно связан с поверхностью эндотелия капилляров. В результате образуются жирные кислоты и глицерин. Часть жирных кислот проходит внутрь жировых клеток, а часть связывается с альбуминами сыворотки крови и уносится с ее током. С током крови может покидать жировую ткань и глицерин.
Расщепление триглицеридов ХМ в печени и в кровеносных капиллярах жировой ткани фактически приводит к прекращению существования ХМ.
Промежуточный обмен липидов . Включает следующие основные процессы: расщепление триглицеридов в тканях с образованием высших жирных кислот и глицерина, мобилизацию жирных кислот из жировых депо и их окисление, образование ацетоновых тел (кетоновых тел), биосинтез высших жирных кислот, триглицеридов, глицерофосфолипидов, сфинголипидов, холестерина и т. д.
Внутриклеточный липолиз
Главным эндогенным источником жирных кислот, используемых в качестве "топлива", служит резервный жир, содержащийся в жировой ткани. Принято считать, что триглицериды жировых депо выполняют в обмене липидов такую же роль, как гликоген печени в обмене углеводов, а высшие жирные кислоты по своей роли напоминают глюкозу, которая образуется в процессе фосфоролиза гликогена. При физической работе и других состояниях организма, требующих повышенной затраты энергии, потребление триглицеридов жировой ткани как энергетического резерва увеличивается.
Поскольку в качестве источников энергии могут использоваться только свободные, т. е. неэтерифицированные, жирные кислоты, то триглицериды сначала гидролизуются при помощи специфических тканевых ферментов - липаз - до глицерина и свободных жирных кислот. Последние из жировых депо могут переходить в плазму крови (мобилизация высших жирных кислот), после чего они используются тканями и органами тела в качестве энергетического материала.
В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют триглицеридлипаза (так называемая гормоночувствительная липаза), диглицеридлипаза и моноглицеридлипаза. Активность двух последних ферментов в 10-100 раз превышает активность первого. Триглицеридлипаза активируется рядом гормонов (например, адреналином, норадреналином, глюкагоном и др.), тогда как диглицеридлипаза и моноглицеридлипаза нечувствительны к их действию. Триглицеридлипаза является регуляторным ферментом.

Установлено, что гормоночувствительная липаза (триглицеридлипаза) находится в жировой ткани в неактивной форме и активируется цАМФ. В результате воздействия гормонов первичный клеточный рецептор модифицирует свою структуру, и в такой форме он способен активировать фермент аденилатциклазу, что в свою очередь стимулирует образование цАМФ из АТФ. Образовавшийся цАМФ активирует фермент протеинкиназу, который путем фосфорилирования неактивной триглицеридлипазы превращает ее в активную форму (рис. 96). Активная триглицеридлипаза расщепляет триглицерид (ТГ) на диглицерид (ДГ) и жирную кислоту (ЖК). Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липолиза - глицерин (ГЛ) и свободные жирные кислоты, которые поступают в кровяное русло.
Связанные с альбуминами плазмы в виде комплекса свободные жирные кислоты с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются либо β-окислению, либо часть их используется на синтез триглицеридов (которые затем идут на образование липопротеидов), глицерофосфолипидов, сфинголипидов и других соединений, а также на этерификацию холестерина.
Другой источник жирных кислот - фосфолипиды мембран. В клетках высших животных непрерывно происходит метаболическое обновление фосфолипидов, в процессе которого образуются свободные жирные кислоты (продукт действия тканевых фосфолипаз).
| Статьи по теме: | |
|
Школьная энциклопедия К какой расе относятся сирийцы
90 % населения Сирии составляют мусульмане, 10% христиане. Мусульмане... Лимонный кекс на кефире с маком
Лимонный кекс на кефире без яиц (с пропиткой) — мой любимый рецепт к... Салат из сайры - простые и оригинальные рецепты аппетитной закуски Салат из сайры консервированной с рисом
Я очень люблю салаты с консервированной рыбой. Их можно готовить... | |